Curcumin Enhances AFM1 Binding to LAB Proteins: A Molecular Docking Study
Curcumin Enhances AFM1 Binding to LAB Proteins: A Molecular Docking Study

Abstract
Aflatoxin M1 (AFM1) is a Group 1 carcinogen and thermostable contaminant of milk and dairy products that resists degradation during conventional processing, including pasteurization and sterilization. Lactic acid bacteria (LAB) provide a biological detoxification strategy through physical adsorption of AFM1 to cell wall components, though the reversibility of this binding limits practical efficacy. Curcumin, a known polyphenol derived from turmeric, has also shown independent AFM1-binding capacity in previous studies. This study investigates the molecular basis for a potential synergistic detoxification mechanism in which curcumin enhances AFM1 binding to LAB surface proteins. Using CB-Dock2, molecular docking was performed with AFM1 and curcumin against two structurally characterized surface proteins from Lactobacillus acidophilus: 4OXD (LdcB LD-carboxypeptidase) and 7QFG (S-layer protein SlpA, domain III). AFM1 bound strongly to 4OXD (ΔG = –8.9 kcal/mol) and moderately to 7QFG (ΔG = –6.9 kcal/mol). Direct curcumin–AFM1 docking yielded a binding affinity of ΔG = –7.7 kcal/mol. In the ternary 4OXD–AFM1–curcumin complex, curcumin occupied a distinct, non-overlapping binding pocket (ΔG = –7.6 kcal/mol) spatially separate from the AFM1 site. These findings provide the first molecular-level evidence that curcumin can bind simultaneously with AFM1 to a LAB peptidoglycan-binding protein without competition, supporting a novel synergistic detoxification model.
1. Introduction
Aflatoxin M1 (AFM1) is a hydroxylated metabolite of aflatoxin B1 (AFB1) produced by Aspergillus flavus and A. parasiticus on contaminated animal feed . Following consumption by lactating mammals, AFB1 is metabolized in the liver and excreted into milk as AFM1, which retains significant toxicity and has been classified as a Group 1 carcinogen by the International Agency for Research on Cancer (IARC) . AFM1 is very stable to heat degradation during pasteurization (73°C) and sterilization (135°C), making it a persistent food safety hazard in the global dairy supply chain , .
Conventional approaches to mitigate AFM1 include physical adsorption using clay-based binders and chemical treatments; however, these methods often compromise the sensory and nutritional quality of milk or raise safety concerns regarding long-term consumption . In response, biological detoxification using food-grade lactic acid bacteria (LAB) has emerged as a promising, safe, and consumer-acceptable alternative. LAB strains, particularly L acidophilus, L. plantarum, L. rhamnosus, and L. casei, have demonstrated significant AFM1-binding capacity in both laboratory media and dairy matrices such as yogurt and kefir, with reported removal efficiencies ranging from 73.9% to 96.9% depending on strain and experimental conditions , , , . The principal mechanism involves physical adsorption to cell wall components, including peptidoglycan and teichoic acids, rather than covalent modification or degradation of the toxin molecule .
A critical limitation of LAB-based AFM1 binding, however, is its reversibility. Under the fluctuating pH conditions of the gastrointestinal tract, up to 60% of bound AFM1 can be released from the bacterial surface, nullifying the intended detoxification benefit , . This observation has motivated the search for complementary agents that could stabilize the AFM1–LAB complex and convert reversible adsorption into permanent sequestration.
Curcumin (diferuloylmethane), the principal bioactive polyphenol of turmeric (Curcuma longa), possesses well-documented antioxidant, anti-inflammatory, and chemopreventive properties , . Recent studies employing density functional theory (DFT) and molecular docking have demonstrated that curcumin forms stable complexes with both AFM1 and AFM2, with adsorption energies of –10.80 kcal/mol for AFM1 at 25°C, primarily through hydrogen bonding and non-covalent interactions . The same study reported that curcumin remained effective at pasteurization temperatures, suggesting practical applicability in dairy processing.
Despite these independent lines of evidence for LAB-mediated adsorption and curcumin–AFM1 binding, the potential for synergistic detoxification, wherein curcumin simultaneously binds to both AFM1 and LAB surface components to enhance overall sequestration, remains largely unexplored. A theoretical review proposed that curcumin could act as a "supramolecular bridge" between AFM1 and bacterial cell wall peptidoglycans, but this hypothesis has not been tested at the molecular level .
The present study uses molecular docking to investigate whether curcumin can bind to LAB surface proteins simultaneously with AFM1, without competing for the same binding pocket. Specifically, it hypothesizes that curcumin occupies a distinct binding site on the 4OXD peptidoglycan-binding protein from L. acidophilus, spatially separated from the AFM1 binding site, thereby supporting a synergistic detoxification model.
2. Research methods and principles
2.1. Protein and ligand structures
Three-dimensional structures of target proteins were obtained from the RCSB Protein Data Bank (PDB) . 4OXD (Structure of the LdcB LD-carboxypeptidase from Lactobacillus acidophilus, resolution 2.8 Å) is a peptidoglycan-binding protein that recognizes the bacterial cell wall and serves as a representative model for LAB surface adsorption sites . 7QFG (Crystal structure of S-layer protein SlpA domain III, residues 309–444, from L. acidophilus, resolution 1.65 Å) represents the crystalline surface layer of the bacterial envelope . Ligand structures, AFM1 (PubChem CID: 15558498) and curcumin (PubChem CID: 969516), were downloaded as 3D SDF files and converted to PDB format before docking.
2.2. Molecular docking
All docking simulations were performed using CB-Dock2 (clab.labshare.cn/cb-dock/), a fully automated online docking server that implements a four-step pipeline including structure preprocessing, cavity detection, and binding affinity scoring . CB-Dock2 treats the uploaded protein as the receptor and searches for potential binding cavities across the entire protein surface, ranking results by binding affinity (ΔG, kcal/mol). Three docking experiments were conducted:
1) AFM1–curcumin (ligand–ligand docking);
2) AFM1–LAB protein (binary binding);
3) Curcumin–AFM1–LAB protein (ternary complex).
For each docking run, the top five cavities were recorded, including binding affinity, cavity volume (ų), and contact residues.
2.3. Data analysis
Binding pockets were compared by examining contact residue lists to determine whether curcumin and AFM1 shared overlapping sets of residues. A non-overlapping pocket was defined as one in which less than 30% of contact residues were shared between the AFM1 and curcumin binding sites.
3. Main results
3.1. Direct curcumin–AFM1 interaction
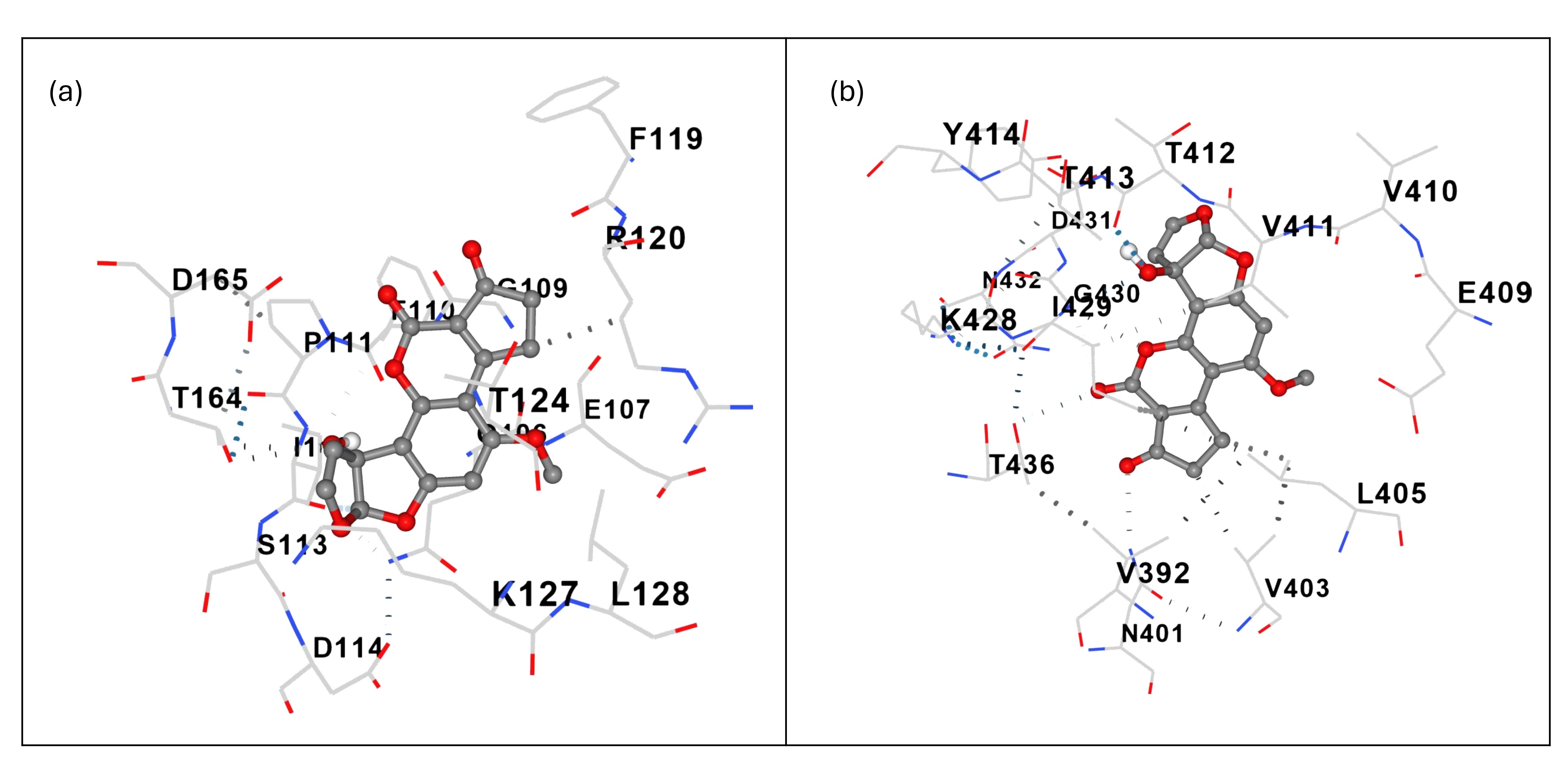
Ligand–ligand docking with AFM1 as the receptor yielded a binding affinity of –7.7 kcal/mol for curcumin (Table 1), indicating a thermodynamically favorable interaction consistent with previous DFT studies (Fig. 1) .
Molecular docking of curcumin to AFM1
3.2. AFM1 binding to LAB proteins
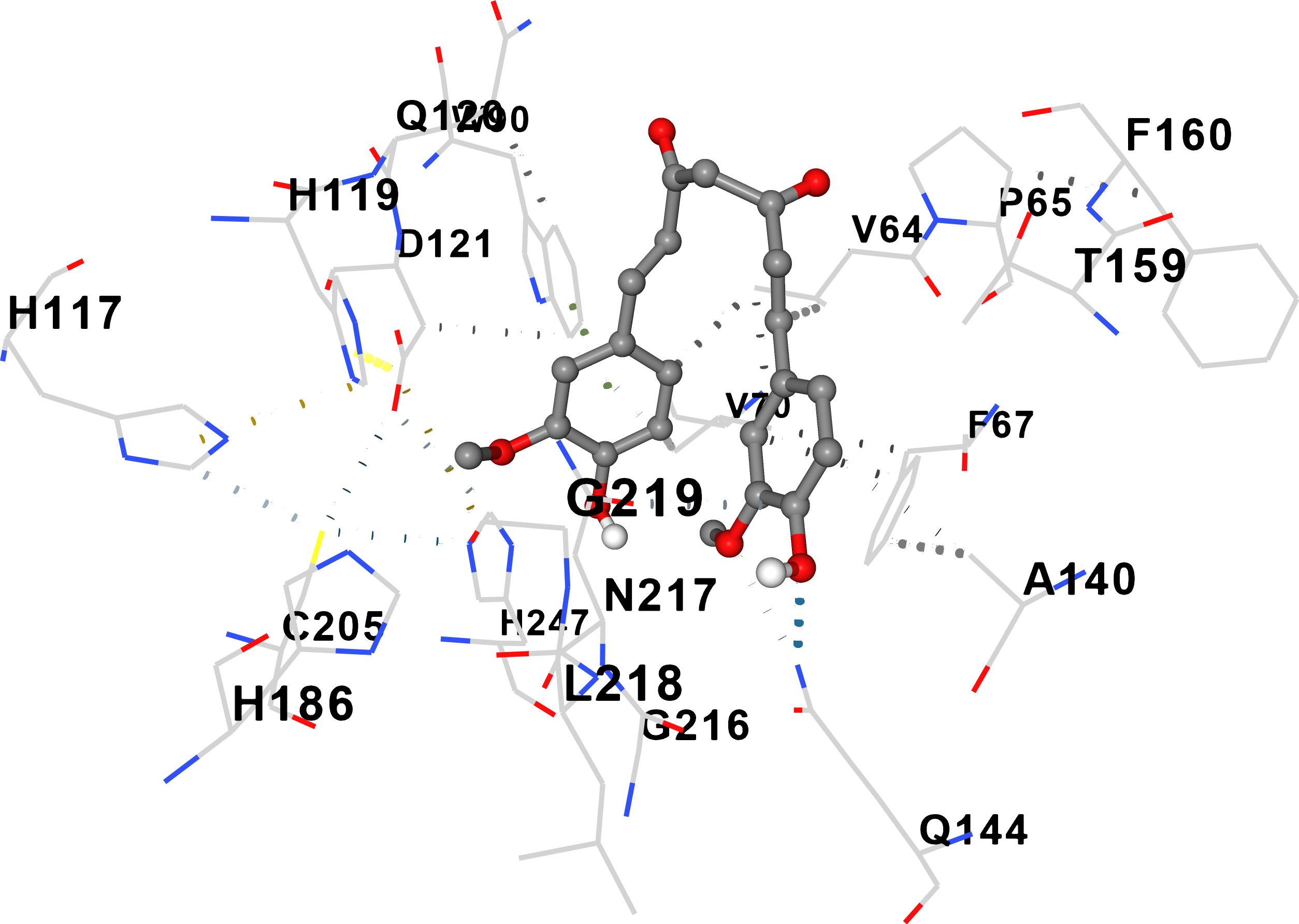
AFM1 docked to 4OXD with a strong binding affinity of –8.9 kcal/mol (cavity 2, volume 3842 ų) (Table 1). Multiple additional cavities exhibited favorable binding (scores –8.3 to –7.1 kcal/mol), suggesting several potential binding loci (Fig. 2a). In contrast, AFM1 binding to 7QFG was moderate (ΔG = –6.9 kcal/mol) with a substantially smaller cavity volume (50 ų), indicating weaker interaction with the S-layer protein (Fig. 2b).
AFM1 binding to Lactobacillus acidophilus surface proteins
Summary of CB-Dock2 docking results for all binding systems
Binding System | Best Cavity | ΔG (kcal/mol) | Volume (ų) | Binding Type |
Curcumin–AFM1 | 1 | –7.7 | 1548 | Direct |
AFM1–4OXD | 2 | –8.9 | 3842 | Binary |
AFM1–7QFG | 4 | –6.9 | 50 | Binary |
Curcumin–4OXD–AFM1 | 1 | –7.6 | 6144 | Non-competitive |
Curcumin–7QFG–AFM1 | 4 | –6.4 | 50 | Competitive |
3.3. Ternary complex analysis
In the preformed 4OXD–AFM1 complex, curcumin bound to cavity 1 with ΔG = –7.6 kcal/mol (volume 6144 ų), which is spatially and compositionally distinct from the AFM1 binding site (Fig. 3).
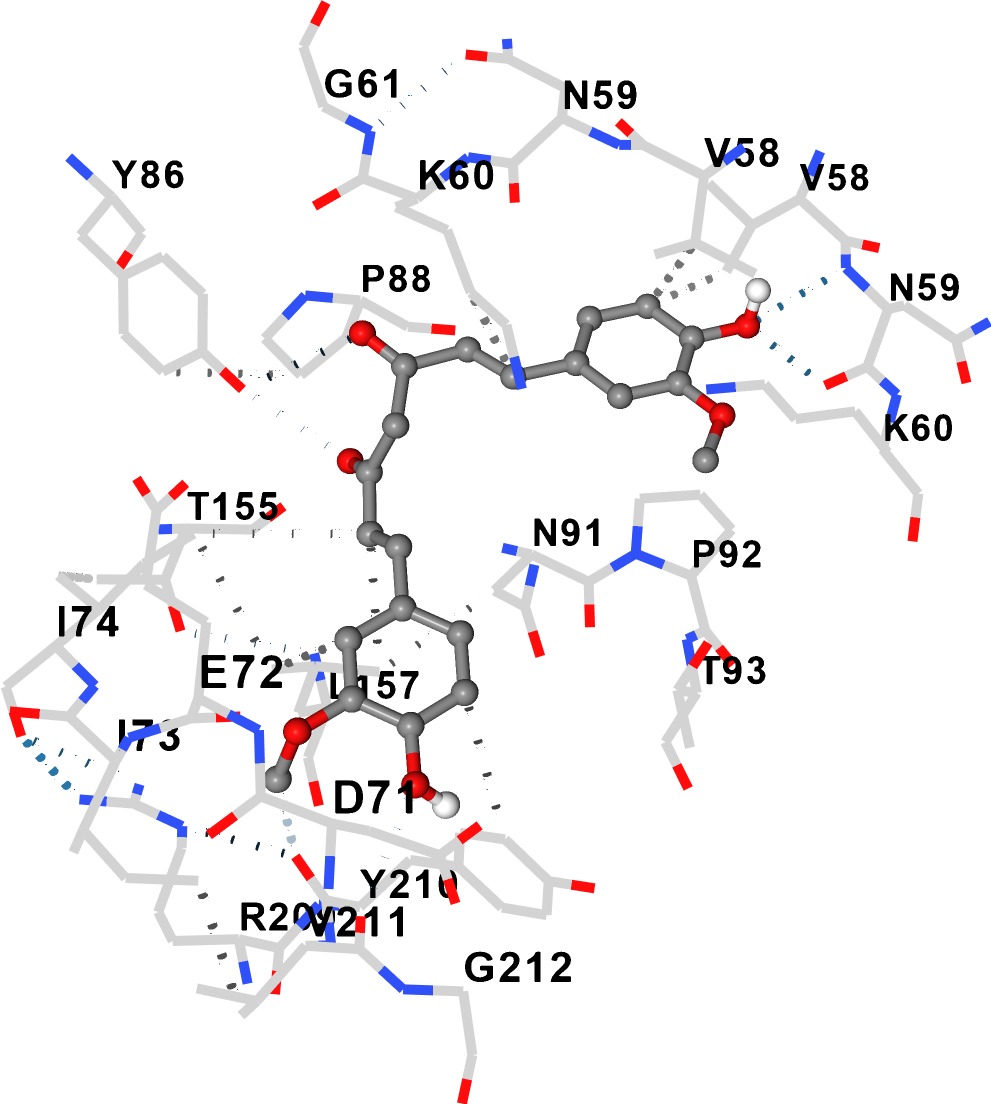
Non-competitive binding of curcumin and AFM1 to 4OXD
Contact residue analysis confirmed no overlap between the two binding pockets (Table 2). By contrast, on 7QFG, curcumin competed for the same pocket as AFM1, with overlapping contact residues indicating competitive binding (Table 2).
Comparative analysis of binding pockets on 4OXD and 7QFG proteins
Protein | Ligand | Key Contact Residues | Pocket Overlap |
4OXD | AFM1 | GLU139:A, ALA140:A, ARG143:A, GLY89:C, GLU90:C, LYS95:C, HIS115:C, TYR116:C | — |
4OXD | Curcumin | TYR63:B, ASP71:B, GLU72:B, ILE73:B, TYR86:B, PRO88:B, GLY89:B, GLU90:B, ARG209:B | None (distinct pockets) |
7QFG | AFM1 | ARG320, ILE321, MET322, VAL392, ASN401, VAL403, VAL404, LEU405, GLU409 | — |
7QFG | Curcumin | ARG320, ILE321, MET322, VAL392, ASN401, VAL403, VAL404, LEU405, GLU409 | Complete (same pocket) |
3.4. Summary of binding affinities
The key findings are:
1) AFM1 binds most strongly to 4OXD (ΔG = –8.9 kcal/mol), substantially more strongly than to 7QFG (ΔG = –6.9 kcal/mol);
2) Curcumin binds directly to AFM1 (ΔG = –7.7 kcal/mol), confirming direct toxin–polyphenol interaction;
3) In the ternary 4OXD system, curcumin binds to a distinct, non-overlapping pocket (ΔG = –7.6 kcal/mol);
4) In the ternary 7QFG system, curcumin competes for the same pocket as AFM1, indicating that synergy is protein-dependent.
4. Discussion
4.1. 4OXD as a superior target for AFM1 binding
The strong binding affinity between AFM1 and 4OXD (ΔG = –8.9 kcal/mol) exceeds typical values reported for small-molecule–protein interactions in aflatoxin detoxification literature. The 4OXD protein, an LdcB LD-carboxypeptidase that recognizes peptidoglycan, represents a functionally relevant model for AFM1 adsorption sites on the LAB cell surface . The large cavity volume and involvement of multiple protein chains suggest that AFM1 inserts deeply into a pre-existing binding groove, forming extensive hydrophobic and polar contacts.
4.2. Distinct binding pockets support a synergistic model
The central finding is that curcumin binds to a different pocket on 4OXD than AFM1. The contact residues for curcumin (cavity 1) are primarily located on chains B and E, whereas the AFM1 binding site (cavity 2) involves chains A, C, and D. This spatial separation provides strong molecular docking evidence that curcumin can bind to the LAB surface simultaneously with AFM1 without steric hindrance. This finding directly supports the "supramolecular bridge" hypothesis proposed in recent theoretical work .
4.3. Implications for binding reversibility
The major limitation of LAB-based AFM1 detoxification is reversibility, with up to 60% of adsorbed AFM1 released under gastrointestinal pH conditions , . By binding to an adjacent pocket, curcumin could exert a stabilizing effect on the overall AFM1–LAB complex through allosteric modulation, formation of a ternary network with additional intermolecular interactions, or physical blockage of the AFM1 exit pathway.
4.4. Protein-dependent competition
The contrasting results for 4OXD (distinct pockets) and 7QFG (competitive binding) underscore that synergistic detoxification potential is protein-specific. S-layer proteins such as 7QFG have smaller binding cavities (42–111 ų) and show competition between AFM1 and curcumin, suggesting they may be less suitable targets for simultaneous binding strategies. LAB strains with abundant peptidoglycan-binding proteins may be superior candidates for curcumin-enhanced detoxification.
4.5. Recommendations for experimental validation
Based on structural insights, the following LAB strains are recommended for experimental validation: Lactobacillus plantarum (high AFM1-binding capacity, up to 80% removal) ; L. rhamnosus (well-characterized for AFM1 binding in yogurt matrices, >50% removal) , ; and L. acidophilus (the species from which 4OXD and 7QFG were derived) . For in vitro validation, incubating selected strains with AFM1-spiked milk in the presence or absence of curcumin is recommended, followed quantifying residual AFM1 by ELISA or HPLC, and measuring AFM1 release after pH washes to assess binding stability.
4.6. Limitations
A few limitations must be noted in this study. Rigid-receptor docking does not account for protein flexibility, which may influence binding pocket geometry. The 4OXD structure is a recombinant domain rather than the intact bacterial cell wall. Therefore, native LAB surface complexity cannot be fully captured by a single protein model. Also, binding energies are computational predictions requiring experimental validation using isothermal titration calorimetry or surface plasmon resonance.
5. Conclusion
This molecular docking study in all likelihood provides the first structural evidence that curcumin can bind simultaneously with AFM1 to a peptidoglycan-binding protein (4OXD) from L. acidophilus without competing for the same binding pocket. AFM1 binds strongly to 4OXD (ΔG = –8.9 kcal/mol) at a site involving chains A, C, and D, while curcumin binds to a distinct, larger cavity (cavity 1, ΔG = –7.6 kcal/mol) primarily involving chains B and E. By contrast, on the S-layer protein 7QFG, curcumin and AFM1 compete for the same shallow pocket. These findings support the idea that curcumin may act as a "supramolecular bridge" stabilizing AFM1–LAB complexes and offer a rational structural basis for developing curcumin-enhanced LAB-based detoxification strategies for dairy products. Experimental validation using candidate strains is now warranted to determine whether curcumin can reduce the reversibility of AFM1 binding and improve dairy product safety.