Evaluation of genetic diversity and genetic structure of Evenk reindeer populations: microsatellite analysis
Evaluation of genetic diversity and genetic structure of Evenk reindeer populations: microsatellite analysis
Abstract
In the Russian Federation, of the four approved breeds of domestic reindeer, the Evenk breed is considered not only the oldest and least studied, but also the least numerous, which increases the risk of genetic isolation in the breed and, as a consequence, changes in the level of genetic diversity, the reduction of which may lead to a narrowing of the population's ability to use internal resources for further development without attracting foreign genetic material. One of the modern approaches to evaluate the state of genetic diversity and population structure is the analysis of microsatellite polymorphism. In the present study, we aimed not only to characterise the state of genetic diversity of the Evenk breed of reindeer bred in the Krasnoyarsk Krai and the Republic of Sakha (Yakutia), but also to assess the genetic structure of the breed in a comparative aspect with other breeds of reindeer and wild reindeer populations. The analysis of the main parameters of allelic and genetic diversity showed that Evenk reindeer of Yakutia are characterised by a higher level of genetic diversity, and also the values of the inbreeding coefficient indicated non-significant deviations of the number of heterozygotes from the theoretically expected. While the sample of Evenk reindeer from the Krasnoyarsk population illustrated an average level of genetic variability. However, it was in this group that the maximum positive values of the inbreeding coefficient were detected, the further accumulation of which may affect local adaptations and co-adapted gene complexes, which will ultimately affect the population's resistance to unfavourable environmental conditions. Cluster analysis identified interesting regularities in the genetic structure of this breed. The Yakutian population of the breed was characterised by the most genetically homogeneous structure, while the Krasnoyarsk population demonstrated the lack of clear genetic consolidation and the presence in its cluster of genetic components of both the cluster of the Evenk breed of Yakutia and the Nenets breed. The results obtained are recommended to be used for the implementation of programmes to preserve, stabilise and increase the breeding stock of the Evenk breed of domestic reindeer.
1. Введение
Северный олень (Rangifer tarandus L., 1758) – единственный представитель рода Rangifer и важный элемент экосистемы Севера . Устойчивость экосистем связана с сохранением, увеличением и использованием биоразнообразия. Сведения о генетической изменчивости и процессах, лежащих в основе происхождения и сохранения видов, играют ключевую роль в понимании структуры и динамики популяций. Поддержание в популяциях оптимальной степени генетической и гетерозиготности необходимо для сохранения способности животных адаптироваться к условиям окружающей среды (изменениям климата, отрицательному действию вредных веществ) Высокое генетическое разнообразие обеспечивает эволюционную приспособленность популяции .
Степень генетической изменчивости между и внутри популяций коррелирует с их демографической историей, а также с факторами окружающей среды . Сведения о генетической структуре популяций способствуют лучшему пониманию, как базовых механизмов эволюции (отбор, мутации, перемещение особей, случайные изменения в геноме) в условиях стресса, так и принятию необходимых мер по восстановлению и рациональному использованию видов . Для корректной оценки состояния генетического разнообразия и структуры популяция актуальным является применение молекулярных методов и подходов молекулярно-генетического анализа . Одним из таких подходов является анализ микросателлитных последовательностей ДНК. Микросателлиты – это широко используемая система маркеров для оценки генетического разнообразия в популяционно-генетических исследованиях, и часто предполагается, что они отражают разнообразие таксона на уровне всего генома . С момента их обнаружения в 1980-х годах, использование микросателлитов линейно росло, и в настоящее время они широко применяются, например, в генетике сохранения видов, судебно-медицинском анализе ДНК, установлении отцовства и исследованиях нейтральной генетической структуры популяций .
В России домашняя популяция северного оленя представлена четырьмя породами, из которых эвенкийская порода считается самой древней и малоизученной из всех известных пород. Ареал обитания этой породы охватывает территорию Красноярского края, Тувы, Бурятии, Якутии (левобережье Лены и южные районы), Хабаровском крае Иркутской, Читинской, Амурской, Сахалинской областей. При этом основное поголовье сосредоточено в Красноярском крае и районах Якутии.
На основании генетической изменчивости микросателлитов исследована генетическая структура региональных популяций ненецкой , чукотской и эвенской пород. Однако исследования генетических особенностей региональных популяций эвенкийской породы ранее не проводились.
Целью настоящей работы являлась оценка уровня генетического разнообразия и генетической структуры эвенкийской породы северного оленя, разводимой на территории Красноярского края и Республики Саха (Якутия) в сравнительном аспекте с другими породами северного оленя, а также популяциями дикого северного оленя, на основании анализа микросателлитов.
2. Материал и методы
В качестве биологического материала были использованы образцы ткани (фрагменты ушной раковины) эвенкийской породы северного оленя, разводимой на территории Красноярского края (EVK_KRA, n = 16) и Республики Саха (Якутия) (EVK_YAK, n = 18). Также в исследования были включены STR-профили пород: ненецкой (NEN, n = 40), эвенской (EVN, n = 33) и чукотской (CHU, n=40). Кроме этого, принимая во внимание факт важной особенности вида – совместное существования домашней и дикой форм , в исследования были включены микросателлитные генотипы диких северных оленей полуострова Таймыра (WLD_T, n=37), районов Якутии (WLD_Y, n=42) и Красноярского края (WLD_KRS, n=18). Таким образом, настоящие исследования включали анализ 256 образцов северных оленей.
Биологический материал и STR-профили были взяты в генетическом банке УНУ «Банк генетического материала домашних и диких видов животных и птицы» в составе сетевой биоресурсной коллекции сБРК СХЖ (соглашение с Минобрнауки России № 075-15-2021-1037 от 28 сентября 2021 г.), созданной и поддерживаемой в ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста. Выделение ДНК проводили с помощью наборов ДНК-Экстран-2 (ЗАО «Синтол», Россия), в соответствии с инструкцией производителя.
Мультиплексный анализ девяти локусов микросателлитов (NVHRT21, NVHRT24, NVHRT76, RT1, RT6, RT7, RT9), проводили на генетическом анализаторе «НАНОФОР 05» (НПК «Синтол», Россия) с последующим определением длин аллелей микросателлитов в программном обеспечении GeneMarker v. 3.0.1. Для обработки полученных результатов формировали матрицу генотипов в программе Microsoft Excel. Анализ внутрипопуляционных параметров включал расчеты показателей аллельного (среднее число аллелей на локус (NА), эффективное число аллелей (NE)) и генетического разнообразия (наблюдаемая (HО) и ожидаемая (HЕ) гетерозиготность и коэффициент инбридинга (FIS)). Генетическую структуру эвенкийской породы северного оленя оценивали с использованием модели Admixture , с использованием смешанной модели (число предполагаемых кластеров, К – от 1 до 10; длина burn-in периода – 100 000; модель Марковских цепей Монте-Карло — 100 000). Определение оптимального количества кластеров (ΔК) для исследованной выборки рыб, было выполнено с использованием приложения STRUCTURE HARVESTER , согласно методу . Статистическую обработку данных проводили с использованием пакета MS Excel 2007 с плагином GenAIEx v. 6.5.1 и программного обеспечения Structure 2.3.4 .
3. Основные результаты и обсуждение
По всей исследуемой выборке северных оленей на основании анализа полиморфизм 9 микросателлитов, было детектировано 111 аллелей. При этом генотипирование животных по 16-ти микросателлитам выявило 106 аллелей на локус . В нашем исследовании, размах вариабельности количества аллелей на локус составил от 9 аллелей в локусе NVHRT24 до 18 аллелей в локусе RT30. Анализ показателей в таблице 1 выявил, что наибольшим уровнем полиморфности характеризовался локус RT1, в котором были выявлены максимальные значения всех показателей, а также отрицательные значения коэффициента инбридинга, свидетельствуя об избытке гетерозиготных вариантов аллелей.
Таблица 1 - Характеристика микросателлитных локусов выявленных в популяциях северных оленей
Локус | Показатель | ||||
NA | NE | HO | HE | FIS | |
NVHRT76 | 6,375±0,596 | 3,383±0,421 | 0,583±0,097 | 0,661±0,052 | 0,126±0,147 |
RT9 | 8,000±0,463 | 4,923±0,492 | 0,610±0,053 | 0,779±0,027 | 0,196±0,102 |
NVHRT24 | 4,625±0,460 | 2,383±0,196 | 0,580±0,066 | 0,551±0,053 | -0,063±0,085 |
RT30 | 9,000±1,018 | 4,510±0,718 | 0,718±0,047 | 0,734±0,042 | 0,024±0,022 |
RT1 | 9,125±1,008 | 5,528±0,782 | 0,770±0,087 | 0,752±0,077 | -0,013±0,045 |
RT6 | 7,750±0,366 | 4,881±0,487 | 0,665±0,036 | 0,775±0,029 | 0,143±0,028 |
RT27 | 7,25±0,559 | 3,996±0,433 | 0,457±0,036 | 0,732±0,024 | 0,380±0,040 |
NV21 | 6,750±0,590 | 4,197±0,308 | 0,456±0,060 | 0,753±0,018 | 0,396±0,079 |
RT7 | 8,375±0,730 | 5,460±0,459 | 0,628±0,067 | 0,807±0,016 | 0,226±0,080 |
Примечание: NA — среднее число аллелей на локус; NE — число эффективных аллелей на локус; HO — наблюдаемая гетерозиготность; HE — ожидаемая гетерозиготность; FIS — коэффициент инбридинга
Наибольшим уровнем полиморфности характеризовался локус RT1, в котором были выявлены максимальные значения всех показателей, а также отрицательные значения коэффициента инбридинга (FIS=0,013), свидетельствуя об избытке гетерозиготных вариантов аллелей. Наименьшие значения показателей среднего и эффективного числа аллелей на локус, а также ожидаемой гетерозиготности были выявлены в локусе NVHRT24. В семи локусах было детектировано уменьшение наблюдаемой гетерозиготности по сравнению с ожидаемой, и как, следствие положительные значения коэффициента инбридинга.
Анализ показателей аллельного разнообразия и степени гетерозиготности (табл. 2), показал, что эвенкийская порода, разводимая на территории Якутии, превосходила все остальные группы по значениям среднего числа аллелей на локус: NA = 8,111 и наблюдаемой гетерозиготности: HO=0,677±0,037.
Таблица 2 - Анализ показателей аллельного разнообразия и степени гетерозиготности популяций эвенкийской породы северного оленя в сравнительном аспекте с породами домашних оленей и популяциями диких северных оленей
Популяция | NA | NE | HO | HЕ | FIS |
CHU | 6,000± 0,527 | 2,929±0,434 | 0,481±0,091 | 0,593±0,062 | 0,146±0,150 |
EVN | 7,222±0,795 | 4,231±0,549 | 0,660±0,058 | 0,735±0,029 | 0,111±0,052 |
EVK_YAK | 8,111±0,633 | 4,079±0,352 | 0,677±0,037 | 0,741±0,021 | 0,084±0,046 |
EVK_KRA | 6,222±0,596 | 4,450±0,482 | 0,563±0,093 | 0,743±0,039 | 0,259±0,122 |
NEN | 8,000±0,645 | 4,564±0,556 | 0,628±0,041 | 0,749±0,041 | 0,161±0,038 |
WLD_T | 8,222±0,619 | 5,062±0,505 | 0,631±0,063 | 0,780±0,029 | 0,196±0,069 |
WLD_Y | 9,778±0,894 | 5,874±0,641 | 0,659±0,044 | 0,807±0,027 | 0,183±0,050 |
WLD_KRS | 6,222±0,521 | 3,710±0,466 | 0,562±0,075 | 0,670±0,066 | 0,116±0,066 |
Примечание: NA — среднее число аллелей на локус; NE — число эффективных аллелей на локус; HO — наблюдаемая гетерозиготность; HE — ожидаемая гетерозиготность; FIS — коэффициент инбридинга. Расшифровку аббревиатур для популяций северного оленя см. в методике
Животные этой породы Красноярского края характеризовались умеренным уровнем, как аллельного, так и генетического разнообразия. Однако, именно в этой группе коэффициент инбридинга имел максимальные положительные значения (FIS=0,259), что указывает на преобладание гомозиготных аллелей.
Значения показателя числа эффективных аллелей на локус, вносящих основной вклад в результат расчета гомо- и гетерозиготности в эвенкиской породе распределились практически равномерно. При этом минимальные значения показателя отмечены в чукотской породе и дикой популяции Красноярского края: NE=2,929±0,434, NE=3,710±0,466 соответственно. Максимальные значения показателя выявлены в двух других популяциях дикого северного оленя: WLD_T =5,062±0,505 и WLD_Y =5,874±0,641. По ожидаемой степени гетерозиготности, исследуемые группы северного оленя имели практически схожие значения. Исключения составила выборка чукотской породы, в которой данный показатель был минимальным: HЕ=0,593±0,062. Коэффициент инбридинга (FIS) во всех выборках, как домашних, так и диких северных оленей, имел положительные значения, отражая преобладание гомозиготных вариантов аллелей. При этом в группе эвенкийских оленей Якутии были выявлены минимальные положительные значения показателя близкие к нулю (FIS=0,084±0,046).
Результаты кластерного анализа представлены на рисунке 1.
Согласно алгоритму, основанному на значениях величины ΔК, было выявлено при К=4 (ΔK = 18,521), значение при К=6 оказались значимыми и более детально отражали генетическую структуру популяций эвенкийской породы северного оленя.
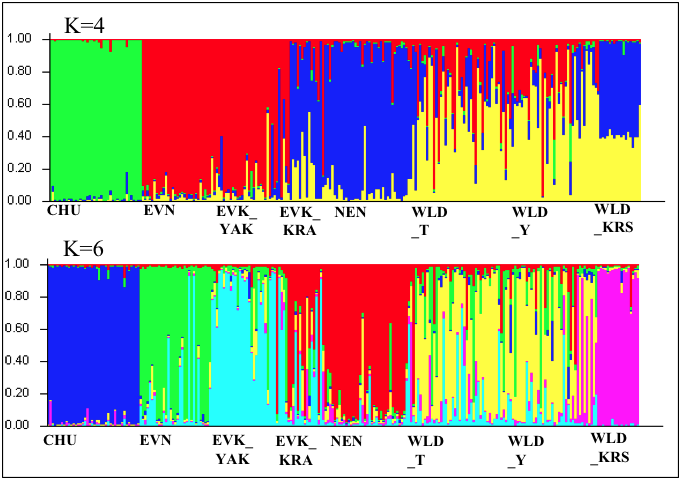
Рисунок 1 - Результаты кластерного анализа эвенкийской породы северного оленя в сравнительном аспекте с породами домашних оленей и популяциями диких северных оленей для числа кластеров К = 4 и K = 6
Примечание: расшифровку аббревиатур для популяций северного оленя см. в методике
4. Заключение
Таким образом, в настоящем исследовании, на основании анализа полиморфизма микросателлитов, нами изучено состояние генетического разнообразия и генетической структуры двух популяций эвенкийской породы северного оленя, разводимой на территории двух регионов: Республики Саха (Якутия) и Красноярского края, в сравнительном аспекте с породами домашнего северного оленя, а также популяциями дикого северного оленя. Полученные результаты выявили, что эвенкийские олени Якутии характеризуются более высоким уровнем генетического разнообразия, а также значения коэффициента инбридинга указывали на недостоверные отклонения количества гетерозигот от теоретически ожидаемого. В то время как выборка северных оленей эвенкийской породы красноярской популяции проиллюстрировала средний уровень генетической изменчивости. Однако именно в этой группе были детектированы максимальные положительные значения коэффициента инбридинга, дальнейшее накопление которого, может оказать влияние на локальные адаптации и коадаптированные генные комплексы, что в конечном итоге, повлияет на устойчивость популяции к неблагоприятным условиям среды. Кластерный анализ выявил интересные закономерности в генетической структуре данной породы. Якутская популяция породы характеризовалась наиболее генетически однородной структурой, в то время как красноярская популяция продемонстрировала отсутствие четкой генетической консолидированности и наличие в своем кластере генетических компонентов, как кластера эвенкийской породы Якутии, так и ненецкой породы. Наблюдаемая интеграция может быть вызвана появлением ненецких и эвенкийских северных оленей Якутии в Красноярском крае для обновления генофонда эвенкийской породы.