CARBOHYDRATE DYNAMICS OF THE GENUS PICEA (РТАСЕАЕ) SPECIES IN THE CONTEXT OF INTRODUCTION

ДИНАМИКА УГЛЕВОДОВ У ПРЕДСТАВИТЕЛЕЙ РОДА PICEA (РТАСЕАЕ) В УСЛОВИЯХ ИНТРОДУКЦИИ
Научная статья
Кищенко И. Т. *
ORCID: 0000-0002-1039-1020,
Петрозаводский государственный университет, Петрозаводск, Россия
* Корреспондирующий автор (ivanki[at]karelia.ru)
АннотацияПри изучении динамики основных углеводов в хвое некоторых интродуцентов рода Picea (в ботаническом саду Петрозаводского университета) установлено, что изменение содержания различных фракций углеводов связано со сменой фенологических фаз. Обнаружены четкие изменения в содержании крахмала, фруктозы и олигосахаридов в период прекращения вегетации и наступления глубокого покоя. Выявлены сходство и рамичия в динамике содержания изучаемых фракций углеводов в хвое аборигенного (Picea abies) и интродуцированных видов рода Picea. Перспективность интродукции многолетних растений, в том числе хвойных, в зоны с довольно суровым климатом во многом определяется степенью их зимостойкости. Подобные реакции растений можно охарактеризовать целым рядом показателей, одним из которых является динамика углеводного обмена в течение года.
Ключевые слова: Picea, интродукция, углеводы, крахмал, глюкоза, фруктоза, дисахариды, олигосахариды.
CARBOHYDRATE DYNAMICS OF THE GENUS PICEA (РТАСЕАЕ) SPECIES IN THE CONTEXT OF INTRODUCTION
Research article
Kishchenko I. T. *
ORCID: 0000-0002-1039-1020,
Petrozavodsk State University, Petrozavodsk, Russia
* Corresponding author (ivanki[at]karelia.ru)
AbstractWhile researching the dynamics of the essential carbohydrates in the needles of some introduced plants of the genus Picea (in the Petrozavodsk University botanical garden), it was concluded that the changes in the amount of various carbohydrate fractions are linked to the phenophase changes. Distinct changes were discovered in the amount of starch, fructose, and oligosaccharides during the completion of the vegetation process and the development of deep dormancy. The similarities and differences in the dynamics of the amount of the researched carbohydrates fractions in the needles of indigenous (Picea abies) and introduced species of the genus Picea were revealed. The perspectives of introducing perennial plants, including the coniferous plants, into harsh climate zones are defined considerably by the extent of their winter resistance. Such reactions can be characterized by a number of features, including the dynamics of carbohydrates metabolism during the year.
Keywords: Picea, introduction, carbohydrates, starch, glucose, fructose, disaccharides, oligosaccharides.
ВведениеИзвестно, что большинство аборигенных видов древесных растений таежной зоны России плохо переносят прогрессирующее загрязнение окружающей среды. Между тем, виды хвойных растений, в том числе и рода Picea других географических районов, устойчивы к загазованности и задымленности, долговечны и весьма декоративны в течение всего года [5], [14]. Кроме того, многие из них отличаются значительно большей продуктивностью, чем местные виды, и нередко способны к натурализации [6], [11]. Повышение биологического разнообразия естественных и искусственных фитоценозов, по мнению ряда исследователей [11], возможно только через интродукцию древесных растений. Все это свидетельствует о необходимости интродукции хвойных и оценки их перспективности. Последняя может быть установлена лишь на основе всестороннего изучения адаптаций, происходящих у испытуемых растений в новых условиях произрастания [4, 1].
В Карелии основным экологическим фактором, лимитирующим рост и развитие растений, является низкая температура воздуха в зимний период. Исследованиями многих авторов [13], [16], [21] показано, что устойчивость растений к низким температурам обеспечивается их подготовкой к периоду глубокого покоя, которая состоит в специальных физиологических и цитологических перестройках. Функции углеводов в древесных растениях многообразны: они являются структурными компонентами клеточных оболочек, резервными защитными соединениями, метаболически активными веществами [19], [3]. Метаболиты представлены моносахаридами, главным образом гексозами: глюкозой и фруктозой. Первым углеводом, образующимся в хлоропластах при фотосинтезе, является фруктозодифосфат, из которого затем формируются другие моносахариды (прежде всего глюкоза), и также ди- и полисахариды [10], [2], [9]. Глюкоза является активным метаболитом, участвует во мномх обменных процессах [8], [3]. Фруктозу можно рассматривать как низкомолекулярный «резерв первой очереди», мобилизуемый при недостатке свободной глюкозы [19] Содержание углеводов в хвое интродуцированных в Карелии видов до сих пор не изучался.
Цель исследований – установить особенности в сезонной динамике содержания углеводов в хвое интродуцированных видов Picea.
Материалы и методы исследований
Исследования проводили в ботаническом саду Петрозаводского государственного университета, расположенного на северном берегу Петрозаводской губы Онежского оз. (средняя подзона тайги, 61 047' с. ш.), в течение 2 лет. Тип почвы — гумусово-железисто-иллювиальный подзол (РН 4.7). Объектами исследований служили представители 6 видов рода Picea: аборигенный вид − ель европейская Picea abies (L.) karst., интродуцированные виды − ель сибирская Picea ovobata Ledeb., ель колючая форма голубая P. pungens Engelm. f. glauca Regel., ель колючая форма зеленая P. pungens Engelm. f. viridis Regel.ель голубая Picea glauca (Moench) Voss., ель сербская Picea omorica (Pane) Purk., ель черная Picea mariana Britt. (см. таблицу 1). Каждый вид представлен 10—25 особями. Условия водного, минерального и светового режима у всех видов одинаковы.
Таблица 1 – Характеристика объектов исследования
| Вид | Место происхождения саженцев (город) | Срдний возраст, лет | Средняя высота, м | Наличие семеношения |
| Piсеа abies Р. obovata Р. рungens f. glauca Р. рungens f. viridis Р. glauca Р. omorica Р. mariana | Петрозаводск Минск С.- Петербург С.- Петербург С.- Петербург Бухарест Бухарест | 31 32 45 45 42 28 28 | 8.4 9.2 16.2 13.7 16.7 5.8 7.1 | + + + + + - - |
Содержание углеводов определяли в однолетней хвое, которую собирали с побегов второго порядка из средней части кроны с разных ее сторон у всех особей каждого вида на 5 стадиях развития: набухания почек (май), интенсивного роста побегов (конец июня), перехода к глубокому покою (середина сентября), глубокого покоя (конец октября), вынужденного покоя (февраль). Содержание углеводов (фруктозы, глюкозы, дисахаридов, олитсахаридов, крахмала) определяли в 4 повторностях по методике, предложенной Г. И. Софроновой с соавт. [18]
Полученные данные оценены на достоверность на 5 %-м уровне значимости.
Результаты и обсуждениеВ хвое большинства изученных видов Picea содержание фруктозы заметно повышается к периоду глубокою покоя (рис. 1). Исключение составляют Р. obovata и Р. mariana. Концентрация глюкозы у Р. abies, Р. рungens f. glauca, Р. рungens f. viridis, Р. glauca и Р. omorica начинает возрастать уже в фазу интенсивного роста побегов, а у остальных видов — только ко времени вхождения в глубокий покой (рис. 1). У Р. mariana сохраняется низкий уровень моносахаридов в разные периоды годичного цикла. Во время вегетации глюкоза и фруктоза активно расходуются на ростовые процессы. По окончании роста снижается использование данных соединений и, кроме того, (наступлением покоя у растений освобождаются моносахариды в результате распад; крахмала. Все это приводит к увеличению количества глюкозы и фруктозы в хвое осенью.
Дисахариды, особенно сахароза, оказывают ингибирующее действие на рост древесных растений. Они повышают устойчивость белков к неблагоприятным воздействиям. Образуя комплексы с белками, дисахариды увеличивают общую водоудерживающую способность клетки и повышают морозостойкость древесного растения [16, 20, 7,]. В хвое Picea всех видов очень невысоко содержание дисахаридов в период вынужденного покоя и во время вегетации, а при переходе к глубокому покою оно резко возрастает. Это происходит, вероятно, в результате гидролиза крахмала, содержание которого с наступлением глубокого покоя снижается (рис.).
Олигосахариды — преобладающая форма углеводов у древесных растений в течение всего года [12]. Они являются ингибиторами роста растения.
Постоянное высокое содержание олиюсахаридов у древесных растений на Севере приводит к замедлению их роста. По данным К. А. Сергеевой [15], особую роль в защите растений от отрицательных температур шрают стахиоза и раффиноза. Образуя комплексные соединения с белками, они повышают устойчивость белковых молекул и общую устойчивость древесных к воздействию отрицательных температур [21].
В хвое всех видов рода Picea при переходе деревьев к глубокому покою отмечается невысокий уровень олигосахаридов, но с наступлением покоя он заметно повышается (рис.). В период вестации у разных видов рода Picea динамика данного класса углеводов неодинакова. У местного вида Р. abies, а также у интродуцированного вида Р. mariana концентрация олигосахаридов меняется незначительно. У Р. рungens f. glauca, Р. рungens f. viridis, Р. glauca и Р. omorica она весьма заметно увеличивается в период интенсивного роста побетв. У Р. obovata очень высокое содержание олигосахаридов отмечено на стадии вынужденного покоя и на начальном этапе роста, а в дальнейшем оно резко снижается. Исследование показало, что изученные виды рода Picea по динамике содержания олигосахаридов в хвое можно условно разделить на 2 группы. К 1-й группе относятся интродуценты, сходные с аборигенным видом (Р. mariana, Р. obovata), а ко 2-й — те, которые значительно отличаются от местното вида (Р. рungens f. glauca, Р. рungens f. viridis, Р. glauca, Р. omorica).
Крахмал в тканях хвойных зимой или отсутствует, или обнаруживается в небольшом количестве. Он накапливается только в период вегетации, т.к. присутствие гранул крахмала в клетках надземных органов зимой резко снижает их морозоустойчивость [13, 17]. Крахмал — основной запасной углевод у древесных растений. Накопление его во время вегетации идет с неодинаковой интенсивностью у разных видов: менее всего оно выражено у Р. omorica, а более вcero — у Р. obovata (рис.). В осенне-зимний период количество крахмала у всех видов очень низкое. К этому времени запасной углевод гидролизуется с образованием растворимых сахаров. У Р. рungens f. glauca и Р. рungens f. viridis отмечена интересная особенность: в период интенсивного роста уровень крахмала невысок, но зато очень велика концентрация глюкозы, дисахаридов, олитсахаридов и выше, чем у других видов, содержание фруктозы. Следовательно, превращение высокомолекулярного углевода в низкомолекулярные растворимые соединения у этого вида происходит уже в июне.
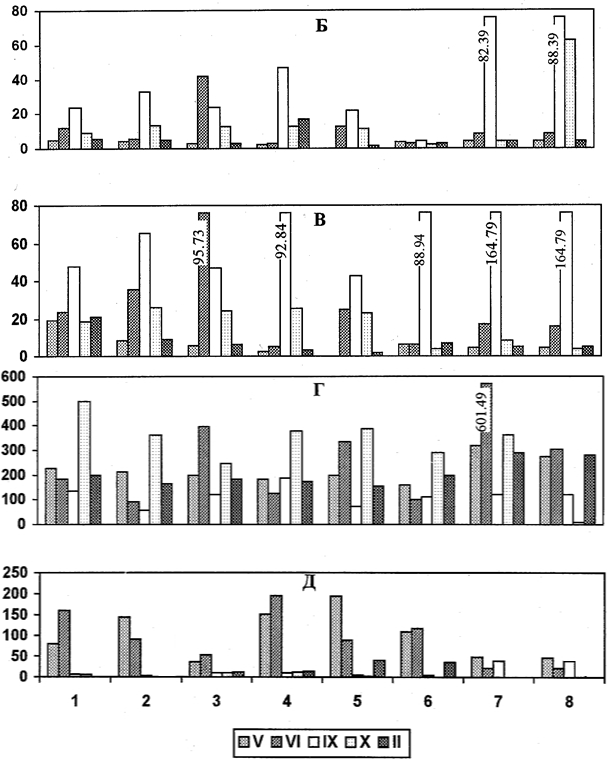
Рис. 1 – Динамика содержания фруктозы (А), глюкозы (Б), дисахаридов (В), олигосахаридов (Г) и крахмала (Д)
Примечание: по оси абсцисс - P. abies (19 лет) - (1), P. abies (50 лет) - (2), P. pungens f. glauca - (3), P. pungens f. viridis - (4), P. glauca - (5), P. mariana -(6), P. omorica - (7), P. obovata - (8). По оси ординат - содержание вещества, мкг/г сыр. вещества
ЗаключениеПроведенные исследования позволили установить, что изменения содержания различных фракций углеводов в хвое видов рода Picea соответствуют смене фенофаз годичного цикла развития растения. Окончание вегетации и переход к состоянию покоя сопровождается гидролизом синтезированного летом крахмала и увеличением в результате этого количества растворимых сахаров: глюкозы и дисахаридов. Наступление глубокого покоя вызывает повышение уровня фруктозы и олигосахаридов. Такая перестройка углеводного обмена направлена на повышение устойчивости растения к низким температурам. Наибольшее значение в процессах развития морозостойкости видов рода Picea среди углеводов имеют олитсахариды. В период глубокого покоя отмечается их высокая концентрация в хвое всех изучаемых видов рода Picea. Динамика олигосахаридов во время роста не одинакова у разных видов Picea. Некоторые интродуценты сходны по данному показателю с аборигенным видом, а другие резко от него отличаются. Выявлены и другие особенности метаболизма углеводов у интродуцентов: у Р. obovata и Р. mariana уровень моносахаридов (глюкозы и фруктозы) намного ниже, чем у местного вида; в то время как у других интродуцентов он намного выше. Кроме того, у Р. omorica, Р. рungens f. glauca и Р. рungens f. viridis в вегетационный период наблюдается наименьшее содержание крахмала, а у Р. obovata ею концентрация максимальна. Низкое содержание этою запасного углевода в хвое Р. рungens f. glauca и Р. рungens f. viridis в период вегетации можно объяснить его распадом до растворимых сахаров, количество которых необычайно высокое.
В целом интродуцированные виды рода Picea, различаясь между собой и с местным видом по соотношению отдельных фракций углеводов, имеют общую стратегию адаптации к перенесению низких температур зимой. Вероятно, поэтому у всех изученных видов рода Picea обмерзание побегов не наблюдалось.
| Конфликт интересов Не указан. | Conflict of Interest None declared. |
Список литературы / References
- Базилевская Н.А. Теория и методы интродукции растений / Н.А. Базилевская. − Москва: Наука, − 1964. − 130 с.
- Веретенников А. В. Физиология растений с основами биохимии / А. В. Веретенников. − Воронеж, − 1987. − 255 с.
- Видершайн Г.Я. Наука об углеводах. Химия и биохимия / Г.Я. Видершайн // Биохимия. − 2009. − Т. 74. − №11. − С. 1582.
- Ворошилов В. Н. Ритм развития у растений / В.Н. Ворошилов − Москва: Наука, − 1960. − 312 с.
- Встовская Т.Н. Интродукция древесных растений дальнего Востока и Западной Сибири / Т.Н. Встовская. − Новосибирск: Наука, −1983. − 196 с.
- Калуцкий К.К. Биоэкологические особенности лесной интродукции / К.К., Калуцкий, Н.А. Болотов // Лесная интродукция. − Воронеж, −1983. − С. 4-
- Красова Н.Г. Динамика накопления содержания углнводов в побегах сортов яблони различной зимостойкости / Н. Г. Красова с соавт. // Плодоводство и ягодоводство России. − 2011. − Т. 28. − №1. − С. 308−315.
- Кретович В.Л. Биохимия растений / В.Л. Кретович. − Москва: Наука, − 1980. − 445 с.
- Кудашева А.В. Динамика накопления углеводов в многолетних злаковых травах Оренбуржья / А.В. Кудашева с соавт. // Вестник Российской академии сельскохозяйственных наук. − 2010. − №2. − С. 51−53.
- Либберт Э. Физиология растений / Э. Либберт. − Москва: Наука, − 1976. − 580 с.
- Мамаев С.А. Проблемы биологического разнообразия и его поддержания в лесных экосистемах / С.А. Мамаев, А.К. Махиев // Лесоведение. − 1996. − № 5.
- Новицкая Ю.Е. Физиологические и биохимические процессы у ели в елово-лиственных насаждениях Севера / Ю.Е. Новицкая // Вопросы селекции, семеноводства и физиологии древесных пород Севера. − Петрозаводск, − 1967. − С. 140—166.
- Новицкая Ю.Е. Особенности физиолого-биохимических процессов в хвое и побегах ели в условиях Севера / Ю.Е. Новицкая. − Ленинград: Наука, − 1971. − 117 с.
- Плотникова Л.С. Научные основы интродукции и охраны древесных растений флоры СССР / Л.С.Плотникова. − Москва: Наука, − 1988. − 263 с.
- Сергеева К. А. О физиологической и биохимической природе глубокого покоя древесных растений / К.А. Сергеева // Симпозиум по физиологии глубокого покоя древесных растений. − Уфа, 1969. − С. 15—17.
- Сергеева К.А. Физиологические и биохимические основы зимостойкости древесных растений / К.А. Сергеева. − Москва: Наука, − 1971. − 174 с.
- Сергеева К.А. Углеводный обмен / К.А. Сергеева // Физиолого-биохимические основы роста и адаптации сосны на Севере. − Ленинград, − 1985. − С. 30− 56.
- Софронова Г.И.. К методике количественного определения углеводов в вегетативных органах сосны обыкновенной / Г.И. Софронова с соавт. // Физиолого-биохимические исследования сосны на Севере. − Петрозаводск, − 1978. − С. 119− 133.
- Судачкова Н.Е. Физиология сосны обыкновенной / Н.Е Судачкова с соавт.. − Новосибирск: Наука, − 1990. − 248 с.
- Тимофеева Г.М. Динамика моно- и олитсахаридов в годичном цикле древесных растений / Г. М. Тимофеева, Л.И. Сергеев // Физиология и биохимия зимостойкости древесных растений. − Уфа, − 1974. − С. 112− 118.
- Туманов И. И. Физиология закаливания и морозостойкости растений / И. И. Туманов. − Москва: Наука, − 1979. − 352 с.
Список литературы на английском языке / References in English
- Bazilevskaya N.A. Teoriya i metody introdukcii rastenij [Theory and methods of plant introduction] / N. A. Bazilevskaya.− Moscow, Science. − 1964. − 130 p. [in Russian]
- Veretennikov A.V. Fiziologiya rastenij s osnovami biohimii [Plant Physiology, the basics of biochemistry] / A.V. Veretennikov. − Voronezh, − 1987. − 255 p. [in Russian]
- Vidershajn G.Ya. Nauka ob uglevodah. Himiya i biohimiya [The Science of carbohydrates. Chemistry and biochemistry] / G.Y. Widersheim // Biohimiya [Biochemistry]. − 2009. – Vol. 74. − No. 11. − P. 15−82. [in Russian]
- Voroshilov V.N. Ritm razvitiya u rastenij [Rhythm of development in plants] / V.N. Voroshilov − Moscow, Science, − 1960. − 312 p. [in Russian]
- Vstovskaya T.N. Introdukciya drevesnyh rastenij dal'nego Vostoka i Zapadnoj Sibiri [Introduction of woody plants of the far East and Western Siberia] / T.N. Vstovskaya. − Novosibirsk: Science, −1983. − 196 p. [in Russian]
- Kaluckij K.K., Bolotov N.A. Bioekologicheskie osobennosti lesnoj introdukcii [Bioecological aspects of forest introduction] / K.K., Kaluski, N.A. Bolotov // Lesnaya introdukciya [Forest introduction]. − Voronezh, −1983. − P. 4−14. [in Russian]
- Krasova N.G. Dinamika nakopleniya soderzhaniya uglnvodov v pobegah sortov yabloni razlichnoj zimostojkosti [Dynamics of accumulation of the content uglevodov in shoots of Apple varieties of different winter hardiness] / N. G. Krasova et al. // Plodovodstvo i yagodovodstvo Rossii [Fruit and berry growing of Russia]. − 2011. – Vol. 28. − No. 1. − P. 308−315. [in Russian]
- Kretovich V.L. Biohimiya rastenij [Biochemistry of plants] / V.L. Kretovich. − Moskva: Nauka, − 1980. − 445 p. [in Russian]
- Kudasheva A.V. Dinamika nakopleniya uglevodov v mnogoletnih zlakovyh travah Orenburzh'ya [Dynamics of carbohydrate storage in perennial cereal grasses of the Orenburg region] / A. V. Kudasheva et al. // Vestnik Rossijskoj akademii sel'skohozyajstvennyh nauk [Herald of the Russian Academy of agricultural Sciences]. − 2010. − No. 2. − P. 51−53. [in Russian]
- Libbert E. Fiziologiya rastenij [Plant Physiology] / Е. Libbert. − Moscow, Science, − 1976. − 580 p. [in Russian]
- Mamaev S.A. Problemy biologicheskogo raznoobraziya i ego podderzhaniya v lesnyh ekosistemah [Problems of biological diversity and its maintenance in forest ecosystems, forestry] / S.A. Mamaev, A.K. Mahiev // Lesovedenie [Forestry]. − 1996. − No. 5. С. 3-10. [in Russian]
- Novickaya Yu.E. Fiziologicheskie i biohimicheskie processy u eli v elovo-listvennyh nasazhdeniyah Severa [Physiological and biochemical processes of Norway spruce in the spruce-deciduous stands in the North] / Y. E. Novitskaya // Voprosy selekcii, semenovodstva i fiziologii drevesnyh porod Severa [Problems of breeding, seed production and physiology of tree species of the North]. − Petrozavodsk, − 1967. − P. 140—166. [in Russian]
- Novickaya Yu.E. Osobennosti fiziolot-biohimicheskih processov v hvoe i pobegah eli v usloviyah Severa [Features fishalot-biochemical processes in the needles and shoots of spruce in the North] / Y.E. Novitskaya. − Leningrad: Science. − 1971. − 117 p. [in Russian]
- Plotnikova L.S. Nauchnye osnovy introdukcii i ohrany drevesnyh rastenij flory SSSR [The Scientific basis of the introduction and protection of woody plants in the flora of the USSR] / L.S. Plotnikova. − Moscow, Science. − 1988. − 263 p. [in Russian]
- Sergeeva K.A. O fiziologicheskoj i biohimicheskoj prirode glubokogo pokoya drevesnyh rastenij [Physiological and biochemical nature of deep rest woody plants] / K.A. Sergeyev // Simpozium po fiziologii glubokoyu pokoya drevesnyh rastenij [Symposium on the physiology of deep dormancy in woody plants]. − Ufa, 1969. − P. 15—17. [in Russian]
- Sergeeva K.A. Fiziologicheskie i biohimicheskie osnovy zimostojkosti drevesnyh rastenij [Physiological and biochemical basis of winter hardiness of woody plants] / К.А. Sergeeva − Moscow, Science. − 1971. − 174 p. [in Russian]
- Sergeeva K.A. Uglevodnyj obmen [Carbohydrate metabolism] Fiziolosh-biohimicheskie osnovy rosta i adaptacii sosny na Severe [Physiolog-biochemical basis of growth adaptation of pine trees in the North] / K.A. Sergeeva. − Leningrad, − 1985. − P. 30− 56. [in Russian]
- Sofronova G.I. K metodike kolichestvennogo opredeleniya uglevodov v vegetativnyh organah sosny obyknovennoj [To the method of quantitative determination of carbohydrates in the vegetative organs of Scots pine] / G.I. Sofronova et al. // Fiziologo-biohimicheskie issledovaniya sosny na Severe [Physiological and biochemical studies of pine in the North]. − Petrozavodsk, − 1978. − P. 119− 133. [in Russian]
- Sudachkova N.E. Fiziologiya sosny obyknovennoj [The physiology of Scots pine] / N.E Sudachkova et al. . − Novosibirsk: Science. − 1990. − 248 p. [in Russian]
- Timofeeva G.M. Dinamika mono- i olitsaharidov v godichnom cikle drevesnyh rastenij [Dynamics of mono- and olitsaccharides in the annual cycle of woody plants] / G.M. Timofeeva, L.I. Sergeev // Fiziologiya i biohimiya zimostojkosti drevesnyh rastenij [Physiology and biochemistry of winter hardiness of woody plants]. − Ufa, − 1974. − P. 112− 118. [in Russian]
- Tumanov I.I. Fiziologiya zakalivaniya i morozostojkosti rastenij [Physiology of hardening and frost resistance of plants] / I.I. Tumanov. − Moscow, Science. − 1979. − 352 p. [in Russian]