THE EFFECT OF BODY SIZE ON FLIGHT PARAMETERS OF APOCRITANS (HYMENOPTERA, APOCRITA)

Фарисенков С.Э.1, Беляев О.А.2, Чуканов В.С.3
1Аспирант, 2соискатель, 3кандидат биологических наук,
Московский государственный университет имени М.В.Ломоносова
Работа выполнена при поддержке Российского научного фонда (№ 14-14-00208)
ВЛИЯНИЕ РАЗМЕРОВ ТЕЛА НА ЛЁТНЫЕ ХАРАКТЕРИСТИКИ СТЕБЕЛЬЧАТОБРЮХИХ ПЕРЕПОНЧАТОКРЫЛЫХ (HYMENOPTERA, POCRITA)
Аннотация
Получены и проанализированы данные по ряду лётных характеристик стебельчатобрюхих перепончатокрылых разного размера. Показано, что с уменьшением размеров тела закономерно изменяются нагрузка на крылья, их удлинение, частота и амплитуда взмахов, сила тяги. Полученные данные помогут понять, какие адаптивные изменения претерпевает полёт в процессе миниатюризации насекомых.
Ключевые слова: размеры тела, полёт, лётные характеристики, Hymenoptera, Apocrita.
Farisenkov S.E.1, Belyaev O.A.2 ,Chukanov V.S.3,
1Postgraduate student, 2postgraduate student, 3PhD in Biology, Lomonosov Moscow State University
The work was supported by Russian Science Foundation(14-14-00208)
THE EFFECT OF BODY SIZE ON FLIGHT PARAMETERS OF APOCRITANS (HYMENOPTERA, APOCRITA)
Abstract
Data on flight parameters of apocritans of different size was obtained and analyzed. It was shown that reduction in body size leads to changes in wing loading and elongation, wingbeat frequency, stroke amplitude, and thrust. Our data could be useful for understanding of adaptive changes of insect flight in the course of miniaturization.
Keywords: body size, flight, Hymenoptera, Apocrita.
Размеры тела во многом определяют характеристики полёта насекомых, наряду с физиологией и морфологическими особенностями строения тела. Характер взаимодействия летящего насекомого с воздушной средой зависит от соотношения сил инерции и сил вязкого трения, выражающегося через число Рейнольдса, диапазон значений которого у насекомых лежит в интервале от 10 до 104 [1]. На примере пчёл было показано, что с уменьшением размеров тела форма крыльев насекомых претерпевает аллометрические изменения [5]. У мельчайших представителей Hymenoptera узкая крыловая пластинка обрамлена длинными щетинками [3, 4], образующими значительную часть его площади, что, по-видимому, является адаптацией к полёту при докритических значениях числа Рейнольдса. В связи с миниатюризацией у стебельчатобрюхих перепончатокрылых отмечается повышенная частота взмахов крыльев [4]. В пределах таксономической группы относительно мелкие насекомые характеризуются более высокой частотой взмахов [1].
В ходе данной работы было исследовано влияние размеров тела различных стебельчатобрюхих перепончатокрылых, в том числе с позиции миниатюризации, на такие характеристики, как нагрузка на крылья, их удлинение, частоту, амплитуду и угол плоскости взмахов крыльев, силу тяги и скорость свободного полёта.
Материалы и методы
Работа выполнена на представителях стебельчатобрюхих перепончатокрылых разных размерных групп и разного систематического положения (Таблица 1). Насекомых отлавливали в летнее время с 2011 по 2015 гг. в окрестностях Ботанического сада МГУ имени М.В.Ломоносова и Звенигородской биологической станции имени С.Н.Скадовского. Эксперименты проводили в лабораторных условиях на живых особях при температуре воздуха 23-27оС.
В лаборатории насекомых помещали в камеру размером 47×13×38 см с прозрачной передней стенкой и осуществляли видеозапись (60 кадров в секунду) свободного прямолинейного полета для последующего вычисления его скорости (V). Для стимуляции насекомых к полёту использовали однонаправленный источник света и обдувание воздухом. Фиксировали максимальные значения скорости полёта для каждой особи.
Затем насекомых обездвиживали с помощью CO2 и крепили за среднеспинку к булавке легкоплавкой смесью воска и канифоли. В закреплённом полёте записывали звук, сопровождающий работу крыльев, для вычисления частоты взмахов (n). Далее снимали высокоскоростное видео (1200 кадров в секунду) с фронтального и бокового ракурсов. Угол плоскости взмахов (β) измеряли в боковой проекции между продольной осью тела и линией, соединяющей крайние положения апекса переднего крыла. За амплитуду взмахов во фронтальной проекции (α’) принимали угол, образованный основанием и крайними положениями апекса переднего крыла. Измерения углов по видеозаписям осуществляли с помощью программы Meazure, вычисление частоты взмахов на основе аудиозаписей – в программе Adobe Audition.
Для измерения движущей составляющей полной аэродинамической силы, или силы тяги (T), булавку с насекомым закрепляли на нижнем конце вертикальной гибкой пластины и фиксировали её максимальное отклонение от положения покоя в миллиметрах при закреплённом полёте. Предварительно пластина была откалибрована грузами: для любого её отклонения с шагом в 1 мм было задано соответствие в ньютонах.
Затем насекомых замаривали и измеряли массу тела (m) на аналитических весах с точностью до 0,1 мг. Длину тела (L) измеряли с помощью окулярной шкалы стереоскопического микроскопа.
Препараты пар крыльев были сфотографированы под микроскопом с апохроматическим объективом. По фотографиям в САПР были измерены длина переднего крыла (l) и суммарная площадь пары крыльев (S).
Был вычислен ряд производных характеристик.
Статистический анализ данных был проведён с помощью программы Statistica. Корреляции между значениями измеренных характеристик оценивали по критерию Пирсона.
Список кратких обозначений измеренных характеристик:
m – масса тела, мг
L – длина тела, мм
l – длина переднего крыла, мм
S – суммарная площадь переднего и заднего крыла, мм2
pw – нагрузка на крылья, Н/м2. ![]() , где g – ускорение свободного падения
, где g – ускорение свободного падения
AR – удлинение крыльев. ![]()
n – частота взмахов крыльев, Гц
V – скорость полета, м/с
T – сила тяги, Н
Tm – относительная сила тяги, Н/кг. ![]()
β – угол плоскости взмахов крыльев, град.
α’ – амплитуда взмахов крыльев во фронтальной проекции, град.
α – амплитуда взмахов крыльев в плоскости взмаха, град. ![]()
Re – число Рейнольдса. ![]() , где ρ – плотность воздуха при 25оС, η – динамическая вязкость воздуха при 25оС
, где ρ – плотность воздуха при 25оС, η – динамическая вязкость воздуха при 25оС
Результаты и обсуждение
Для исследованных видов были получены значения большинства измеряемых характеристик. Оригинальные данные были частично дополнены почерпнутыми из литературы измерениями [12] в тех случаях, когда их оказалось невозможно получить самостоятельно. Медианные значения характеристик для каждого вида представлены в таблице 1.
Таблица 1 – медианные значения измеренных характеристик
(* – по Вейс-Фо [12])
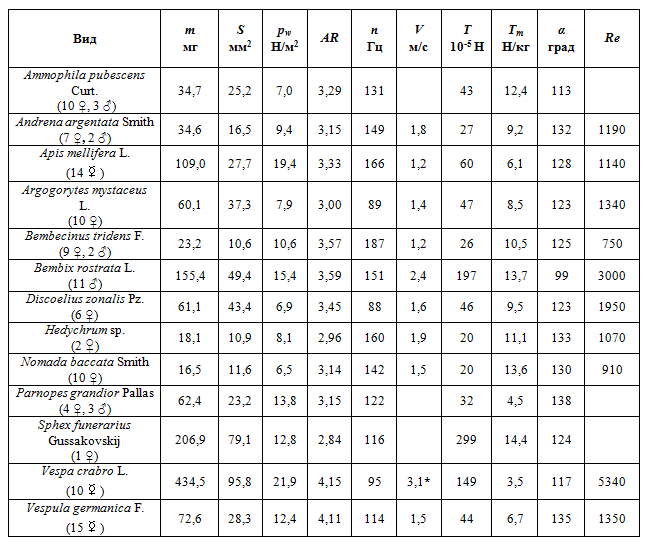
Минимальная масса тела отмечена у Nomada baccata (11,8 мг), максимальная у Vespa crabro (500 мг). Значения числа Рейнольдса у исследованных особей находится в диапазоне от 430 (Bembecinus tridens) до 5500 (Vespa crabro).
Площадь крыльев нелинейно убывает при уменьшении массы тела. Зависимость имеет вид ![]() , k ≈ 2,2 (рис. 1а).
, k ≈ 2,2 (рис. 1а).

Рис. 1 – Графики зависимостей:
а) площади крыльев от массы тела, б) удлинения крыльев от массы тела, в) частоты взмахов от массы тела, г) частоты взмахов от массы тела и площади крыльев, д) относительной силы тяги от массы тела, е) амплитуды взмахов от площади крыльев. Точками обозначены измерения, полученные для всех исследованных особей
Нагрузка на крылья изменяется в зависимости от массы тела согласно функции ![]() , k ≈ 2,2. Значения нагрузки на крылья лежат в интервале от 3,9 (Bembecinus tridens) до 29,7 (Apis mellifera).
, k ≈ 2,2. Значения нагрузки на крылья лежат в интервале от 3,9 (Bembecinus tridens) до 29,7 (Apis mellifera).
Удлинение крыльев уменьшается с понижением массы тела (рис. 1б), что было ожидаемо, исходя из данных литературы [4, 5]. Характер зависимости удлинения от массы тела для данной выборки выявить достаточно сложно в силу большой дисперсии значений удлинения крыльев. Поскольку удлинение крыльев является адаптивной характеристикой и связано с оптимизацией формы крыла к полёту, на него могут влиять не только масса тела, но и многие другие морфологические и физиологические характеристики, присущие конкретному виду. Удлинение связано с величиной индуктивного сопротивления крыла относительно величины его общего лобового сопротивления, что в свою очередь зависит от числа Рейнольдса [5]. Действительно, нами было обнаружено, что удлинение крыла положительно коррелирует с числом Рейнольдса.
Перепончатокрылым меньшего размера свойственна более высокая частота взмахов крыльев (рис. 1в): показана отрицательная корреляция частоты взмахов крыльев с массой тела и числом Рейнольдса. Характер зависимости частоты взмахов от массы тела, по-видимому, носит степенной характер вида ![]() [11]. Однако гораздо более точно частоту взмахов определяет формула
[11]. Однако гораздо более точно частоту взмахов определяет формула ![]() , учитывающая также площадь крыльев [6]. При применении её к полученным данным мы получили близкое к линейному распределение (рис. 1г), которое аппроксимируется функцией:
, учитывающая также площадь крыльев [6]. При применении её к полученным данным мы получили близкое к линейному распределение (рис. 1г), которое аппроксимируется функцией:
![]() , где
, где ![]() , k ≈ 360.
, k ≈ 360.
Из этого следует, что частота взмахов у перепончатокрылых во многом определена размерами тела, а именно его массой и площадью крыльев.
Скорость полета в экспериментальных условиях не коррелирует ни с какими характеристиками, кроме числа Рейнольдса. Такой результат является следствием большой дисперсии значений скорости полёта в пределах даже одного вида среди исследованных насекомых и может быть связан с их различным физиологическим состоянием на момент проведения эксперимента. Для получения достоверных средних значений скорости полёта и статистически значимых корреляций необходимо большее число измерений.
Сила тяги нелинейно зависит от массы тела, характер зависимости имеет вид ![]() , где k ≈ 3,5. Соответственно с уменьшением массы тела относительная сила тяги увеличивается согласно функции
, где k ≈ 3,5. Соответственно с уменьшением массы тела относительная сила тяги увеличивается согласно функции ![]() , k ≈ 29 (рис. 1д). Описанную зависимость можно объяснить тем, что удельная по массе сила летательных мышц животных пропорциональна массе тела в степени -1/3 [7]. Кроме того, у мелких насекомых сила тяги превалирует над подъёмной силой, так как с уменьшением числа Рейнольдса возрастает относительная величина сил вязкого трения, и, как следствие, увеличивается соотношение лобового сопротивления к массе тела [9]. Чтобы выяснить степень влияния перечисленных факторов, необходимы измерения подъёмной силы у исследованных видов. Стоит отметить, что Bembix rostrata демонстрируют необычайно высокую силу тяги для своей массы тела, что, по всей видимости, имеет для них адаптивное значение, так как для прокорма потомства они отлавливают быстро и маневренно летающих двукрылых (слепней, журчалок и др.) [2, 10].
, k ≈ 29 (рис. 1д). Описанную зависимость можно объяснить тем, что удельная по массе сила летательных мышц животных пропорциональна массе тела в степени -1/3 [7]. Кроме того, у мелких насекомых сила тяги превалирует над подъёмной силой, так как с уменьшением числа Рейнольдса возрастает относительная величина сил вязкого трения, и, как следствие, увеличивается соотношение лобового сопротивления к массе тела [9]. Чтобы выяснить степень влияния перечисленных факторов, необходимы измерения подъёмной силы у исследованных видов. Стоит отметить, что Bembix rostrata демонстрируют необычайно высокую силу тяги для своей массы тела, что, по всей видимости, имеет для них адаптивное значение, так как для прокорма потомства они отлавливают быстро и маневренно летающих двукрылых (слепней, журчалок и др.) [2, 10].
Выявлена обратная зависимость амплитуды взмахов от массы тела, числа Рейнольдса, а также площади и длины крыльев (рис. 1е). При уменьшении размеров крыла, его момент инерции уменьшается нелинейно, пропорционально квадрату длины, поэтому крылья мелких Hymenoptera обладают относительно меньшей инерционностью [8], что позволяет им совершать взмахи с большей амплитудой и частотой. Из общей зависимости выбиваются Vespa crabro со сравнительно большой амплитудой взмахов.
Заключение
Размер тела оказывает значительное влияние на форму крыльев и лётные характеристики исследованных насекомых. Мелкие стебельчатобрюхие перепончатокрылые имеют низкую нагрузку на крылья и меньшее удлинение крыльев. Они обладают большей частотой и амплитудой взмахов по сравнению с крупными представителями подотряда, которые уступают им так же по показателю относительной силы тяги.
Благодарности
Авторы глубоко признательны Антропову Александру Валентиновичу, Левченко Тимофею Викторовичу, Карцеву Владимиру Михайловичу, Винокурову Николаю Борисовичу, Лютиковой Ларисе Ивановне, Гаврилову Валерию Михайловичу, Жантиеву Рустему Девлетовичу, Полилову Алексею Алексеевичу.
Литература
- Бродский А.К. Механика полёта насекомых и эволюция их крылового аппарата. – Л.: Изд-во Ленинградского университета, 1988. – 208 с.
- Длусский Г.М. Отряд Перепончатокрылые. В кн.: Жизнь животных. Т. 3. – 2-е изд. – М.: Просвещение, 1984. – С. 339-388.
- Прингл Дж. Полёт насекомых. – М.: Изд-во иностранной литературы, 1963. – 180 с.
- РасницынА.П. Происхождение и эволюция перепончатокрылых насекомых. – М.: Наука, 1980. – 192 с.
- Danforth B.N. The evolution of hymenopteran wings: the importance of size // J. Zool., Lond. 1989. 218. P. 247-276.
- Deakin M. Formulae for insect wingbeat frequency // Journal of Insect Science. 2008. Vol. 10. № 96. P. 1-9.
- Ellington C.P. Limitations on animal flight performance // Exp. Biol. 1991. Vol. 160. P. 71-91.
- Greenewalt, C.H. The wings of insects and birds as mechanical oscillators // Proc. Amer. Philosoph. Soc. 1960. 104. № 6. P. 605-611
- Horridge G.A. The flight of very small insects // Nature. 1956. Vol. 178. P. 1334-1335.
- New T.R. Hymenoptera and Conservation. – John Wiley & Sons. 2012. – 232 pp.
- Sudo S. et al. The wing apparatus and flapping behavior of Hymenoptera // JSME International Journal. Series C. 2001. Vol.44. № 4. P. 1103-1110.
- Weis-Fogh T. Respiration and tracheal ventilation in locust and other flying insects // J. Exp. Biol. 1967. Vol. 47. P. 561-587.
References
- Brodskij A.K. Mekhanika poljota nasekomyh i evoljutsija ih krylovogo apparata – : Izd-vo Leningradskogo universiteta, 1988. – 208 s.
- Dlusskij G.M. Otryad Pereponchatokrylye. V kn.: Zhizn zhivotnyh. T. 3. – 2-e izd. – M.: Prosveshchenie, 1984. – S. 339-388.
- Pringle J. Poljot – M.: Izd-vo innostrannoj literatury, 1963. – 180 s.
- RasnitsynP. Proishozhdenie i evoljutsija pereponchatokrylyh nasekomyh. – M.: Nauka, 1980. – 192 s.
- Danforth B.N. The evolution of hymenopteran wings: the importance of size // J. Zool., Lond. 1989. 218. P. 247-276.
- Deakin M. Formulae for insect wingbeat frequency // Journal of Insect Science. 2008. Vol. 10. № 96. P. 1-9.
- Ellington C.P. Limitations on animal flight performance // Exp. Biol. 1991. Vol. 160. P. 71-91.
- Greenewalt, C.H. Thewings of insects and birds as mechanical oscillators // Proc. Amer. Philosoph. Soc. 1960. 104. № 6. P. 605-611
- Horridge G.A. The flight of very small insects // Nature. 1956. Vol. 178. P. 1334-1335.
- New T.R. Hymenoptera and Conservation. – John Wiley & Sons. 2012. – 232 pp.
- Sudo S. et al. The wing apparatus and flapping behavior of Hymenoptera // JSME International Journal. Series C. 2001. Vol.44. № 4. P. 1103-1110.
- Weis-Fogh T. Respiration and tracheal ventilation in locust and other flying insects // J. Exp. Biol. 1967. Vol. 47. P. 561-587.