ИССЛЕДОВАНИЕ ОТСТАВЛЕННЫХ ЭФФЕКТОВ ДЕЙСТВИЯ ОСТРОЙ НОРМОБАРИЧЕСКОЙ ГИПОКСИИ НА РАЗВИТИЕ КРЫСЯТ В ТЕСТАХ «ПРОСТРАНСТВЕННОЕ ОБУЧЕНИЕ В ЛАБИРИНТЕ «БАРНС»» И «СЛОЖНЫЙ ПИЩЕВОЙ ЛАБИРИНТ»
ИССЛЕДОВАНИЕ ОТСТАВЛЕННЫХ ЭФФЕКТОВ ДЕЙСТВИЯ ОСТРОЙ НОРМОБАРИЧЕСКОЙ ГИПОКСИИ НА РАЗВИТИЕ КРЫСЯТ В ТЕСТАХ «ПРОСТРАНСТВЕННОЕ ОБУЧЕНИЕ В ЛАБИРИНТЕ «БАРНС»» И «СЛОЖНЫЙ ПИЩЕВОЙ ЛАБИРИНТ»
Аннотация
Перинатальная гипоксия является значимой проблемой, оказывающей влияние на развитие нервной системы и последующие когнитивные функции как у человека, так и у животных. Цель — изучение отставленных эффектов действия острой нормобарической гипоксии на развитие крысят в тестах «Пространственное обучение в лабиринте «Барнс» и «Сложный пищевой лабиринт». Работа проводилась на крысах обоего пола линии Wistar (20 половозрелых самок и 10 половозрелых самцов, которые поровну по принципу рандомизации были разделены на 2 группы и от них было получено потомство). 1 группа — контроль, а животным 2 группы моделировали состояние недоношенной беременности человека: воздействие острой нормобарической гипоксии на крысят на вторые постнатальные сутки и впоследствии оценивали физическое развитие в адолесцентном возрасте. Установлено, что острая нормобарическая гипоксия приводит к расстройствам поведенческих реакций у крыс адолесцентного возраста, что в тесте «Пространственное обучение в лабиринте «Барнс» выражается дефектами в обучении и памяти, а в тесте «Сложный пищевой лабиринт» — нарушением выработки пищедобывательного рефлекса на место.
1. Введение
Перинатальная гипоксия является значимой проблемой, оказывающей влияние на развитие нервной системы и последующие когнитивные функции как у человека, так и у животных. Эти состояния, возникающие в период внутриутробного развития или непосредственно перед и после рождения, могут приводить к недостатку кислорода в тканях, что особенно критично для формирующегося мозга
, , .У человека перинатальная гипоксия может быть связана с различными неврологическими и психическими расстройствами. Исследования показывают, что дети, перенесшие гипоксию в этом периоде, чаще сталкиваются с проблемами в обучении, снижением памяти, задержками в развитии и трудностями с вниманием. Эти нарушения могут быть связаны с недостаточным развитием нейронных связей в ключевых областях мозга, таких как гиппокамп, отвечающем за обработку памяти и обучение
, , .В экспериментах на животных, например, на моделях крыс, перинатальная гипоксия демонстрирует аналогичные паттерны. У крысят, подвергшихся гипоксии, отмечено снижение объема гиппокампа и нарушенная синаптическая пластичность, что критично для формирования долговременной памяти. Такие животные проявляют трудности в выполнении задач, требующих обучения, и часто отличаются повышенной тревожностью и нарушением поведения, что также может негативно сказываться на познавательных процессах
, .Механизмы, лежащие в основе нарушений памяти и способности к обучению при перинатальной гипоксии, могут включать изменения в нейрохимических путях, нарушении нейрогенеза и перераспределении ресурсов мозга. Исследования показывают, что снижение уровня нейротрофических факторов и серотонина может ограничивать возможности адаптации и выживания нейронов, что, в свою очередь, влияет на когнитивные функции
, .Таким образом, перинатальная гипоксия представляет собой серьезный риск для формирования памяти и способности к обучению, как у людей, так и у животных. Эти последствия могут оказывать длительное влияние на качество жизни и требуют внимательного подхода к их профилактике и реабилитации в ранние периоды развития.
Цель — изучение отставленных эффектов действия острой нормобарической гипоксии на развитие крысят в тестах «Пространственное обучение в лабиринте «Барнс» и «Сложный пищевой лабиринт».
2. Методы и принципы исследования
Работа проводилась на крысах обоего пола линии Wistar (20 половозрелых самок и 10 половозрелых самцов, которые поровну по принципу рандомизации были разделены на 2 группы). Животных содержали в стандартных условиях вивария. От взрослых крыс были получены крысята и каждый выводок содержался в отдельной клетке вместе с матерью до достижения месячного возраста, а затем крысят отсаживали от матери. В первой группе от них было получено 122 крысенка (69 самцов и 53 самки), а во второй 128 крысят (59 самцов и 69 самок).
Исследование выполнено на модели недоношенной беременности человека: воздействие острой нормобарической гипоксии (ОНГ) на крысят на вторые постнатальные сутки в инкубаторе New Brunswick Scientific «Innova CO-48 CO2» (США) .
День рождения крысят принимали за нулевой день жизни. На второй день жизни животных делили на равные группы:
1 группа — контроль, крысята обоих полов, не подвергавшиеся действию гипоксии, но на второй день жизни они находились в инкубаторе при 37 ºС в течение 2-х часов, при атмосферном содержании кислорода в воздухе.
2 группа — экспериментальная, крысята обоих полов подергались действию острой нормобарической гипоксии, путем помещения в инкубатор с пониженным содержанием кислорода (8% О2) и температурой 37 ºС на 2 часа.
Для исследования поведения и развития памяти и способности к обучению у крыс, перенесших гипоксию на вторые сутки жизни, проводили тесты на пространственное обучение в лабиринте Барнса — на 42–47 день жизни и на выработку условного пищедобывательного рефлекса на место («Сложный пищевой лабиринт») — на 42–47 день жизни.
Тест «Пространственное обучение в лабиринте «Барнс» позволяет оценить пластичность обучения, регулируемую гиппокампом. Лабиринт «Барнс» представляет собой круглую арену диаметром 122 см, расположенную на высоте 113 см, на которой находится 18 лунок, расположенных по периферии арены на равном удалении друг от друга. 17 лунок представляют собой небольшие углубления диаметром 9,5 см — ложные норки, одна из лунок соединена со съемным большим убежищем размером 11×30 см, находящимся под ареной — истинная норка. Сама установка расположена в изолированном пространстве, окружена тремя стенами и ширмой (зона тестирования), на одной из стен расположен визуальный ориентир. В качестве стрессогенного стимула использовалось равномерное яркое освещение арены (900 лк). При обучении каждой крысы выбиралось индивидуальное расположение истинной норки, которое сохранялось на протяжении всех дней эксперимента. Между животными расположение истинной норки менялось по следующей схеме: 1,10, 2, 11, 3, 12, и т. д.
Перед началом эксперимента клетки с животными переносили из комнаты содержания в зону тестирования не менее чем за 30 минут до начала проведения тестирования. Крысу помещали в непрозрачный цилиндр с крышкой, находящийся в центре арены лабиринта на 10–15 секунд во избежание ориентации животного до начала эксперимента. Начало попытки отсчитывалось с момента поднятия цилиндра. Максимальное время поиска истинной норки для каждой попытки составляло 3 минуты. Перерыв между попытками составлял около 2-х минут, после каждой попытки арена и норки протиралась 10 % раствором этанола.
В нашей схеме эксперимента животных обучали в течение 5 дней по 4 попытки/день. На 6 день в ходе 2-х попыток проводили проверку воспроизведения навыка в условиях высокой стрессогенной нагрузки (электрический звонок, создающий звук интенсивностью 100 дБ). В каждую попытку оценивали следующие параметры: время нахождения истинной норки и количество ошибок (заглядываний, обнюхиваний ложных норок).
«Сложный пищевой лабиринт» представляет собой квадратную камеру, разделенную пятью прозрачными перегородками на 6 коридоров. В каждой перегородке присутствует прямоугольное отверстие. Перед экспериментом животных подвергали 24-часовой пищевой депривации.
1. Адаптация. В первый день эксперимента крыс помещали в лабиринт на 30 минут с целью адаптации и угашения ориентировочно-исследовательской реакции.
2. Обучение. В последующие 4 дня животное помещали в лабиринт по 5 раз подряд ежедневно, при этом длительность каждой посадки не превышала 3-х минут. Животное помещалось в стартовый отсек, после чего визуально регистрировалось следующие показатели:
- число выполненных реакций (число случаев, когда животное находит пищевое подкрепление в течение 3-х минут пребывания в лабиринте);
- количество ошибок (число любых отклонений от правильной траектории движения).
В дни опыта животных кормили один раз в сутки непосредственно после эксперимента.
Цифровой материал всех экспериментов подвергали статистической обработке с помощью пакета программ STATISTICA Application 10.0.1011.0. Оценку нормальности распределения данных проводили с помощью критерия Шапиро-Уилка. Так как характер распределения данных соответствовал нормальному результату эксперимента оценивали с помощью методов дисперсионного анализа (ANOVA/MANOVA). В случае достоверного влияния выбранных факторов или их взаимодействия, проводили последующий post hoc анализ с использованием критерия Фишера (Fisher LSD test). Межгрупповые различия динамических показателей (прибавка веса) оценивались с помощью ANOVA для повторных измерений. На рисунках данные представлены в виде среднего значения и ошибки среднего арифметического.
3. Основные результаты
Результаты теста «Пространственное обучение в лабиринте «Барнс» представлены на рисунках 1 и 2.
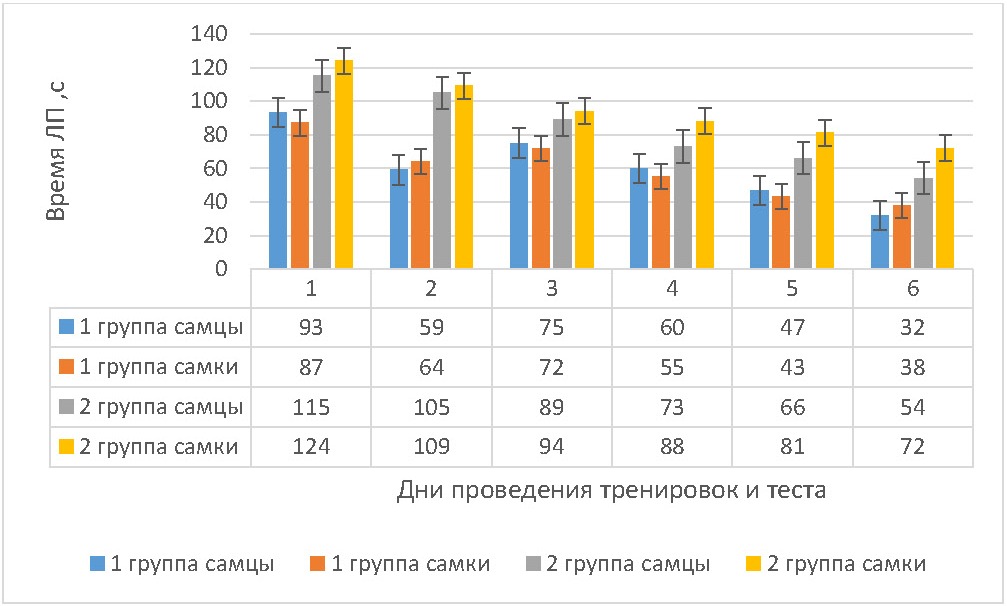
Рисунок 1 - Динамика времени латентного периода захода в истинные норки в дни тренировок
Примечание: M±m
У самцов 2 группы время ЛП на 2 день тренировок снизилось на 8,7%, на 3 день — на 22,6% (p <0,001), на 4 день — на 36,5% (p <0,028), на 5 день — на 42,6% (p <0,034) и на 6 день — на 53,0% (p <0,001) от первоначального значения. У самок 2 группы время ЛП на 2 день тренировок снизилось на 12,1% (p <0,017), на 3 день — на 24,2% (p <0,037), на 4 день — на 29,0% (p <0,001), на 5 день – на 34,7 % (p <0,001) и на 6 день – на 41,9 % (p <0,023) от первоначального значения. При этом различия во времени ЛП с учетом гендерного фактора во 2 группе животных были достоверным на 4 день тренировок (у самок больше, чем у самцов на 20,6% (p <0,001)), на 5 день тренировок (у самок больше, чем у самцов на 22,7% (p <0,003)) и на 6 день теста (у самок больше, чем у самцов на 33,3% (p <0,021)).
При этом время латентного периода захода в истинные норки у самцов и самок крыс 2 группы было больше, чем у крыс 1 группы соответствующего пола: у самцов 2 группы в 1 день время ЛП было больше на 23,7% (p <0,011), во 2 день — на 77,8% (p <0,024), в 3 день — на 18,7% (p <0,037), в 4 день — на 21,7% (p <0,029), в 5 день — на 40,4% (p <0,001) и в 6 день — на 68,8% (p <0,001) по сравнению с самцами 1 группы; у самок 2 группы в 1 день время ЛП было больше на 42,5% (p <0,021), во 2 день — на 70,3% (p <0,033), в 3 день — на 30,6% (p <0,041), в 4 день — на 60,0% (p <0,037), в 5 день — на 88,4% (p <0,001) и в 6 день — на 89,5% (p <0,001) по сравнению с самками 1 группы.
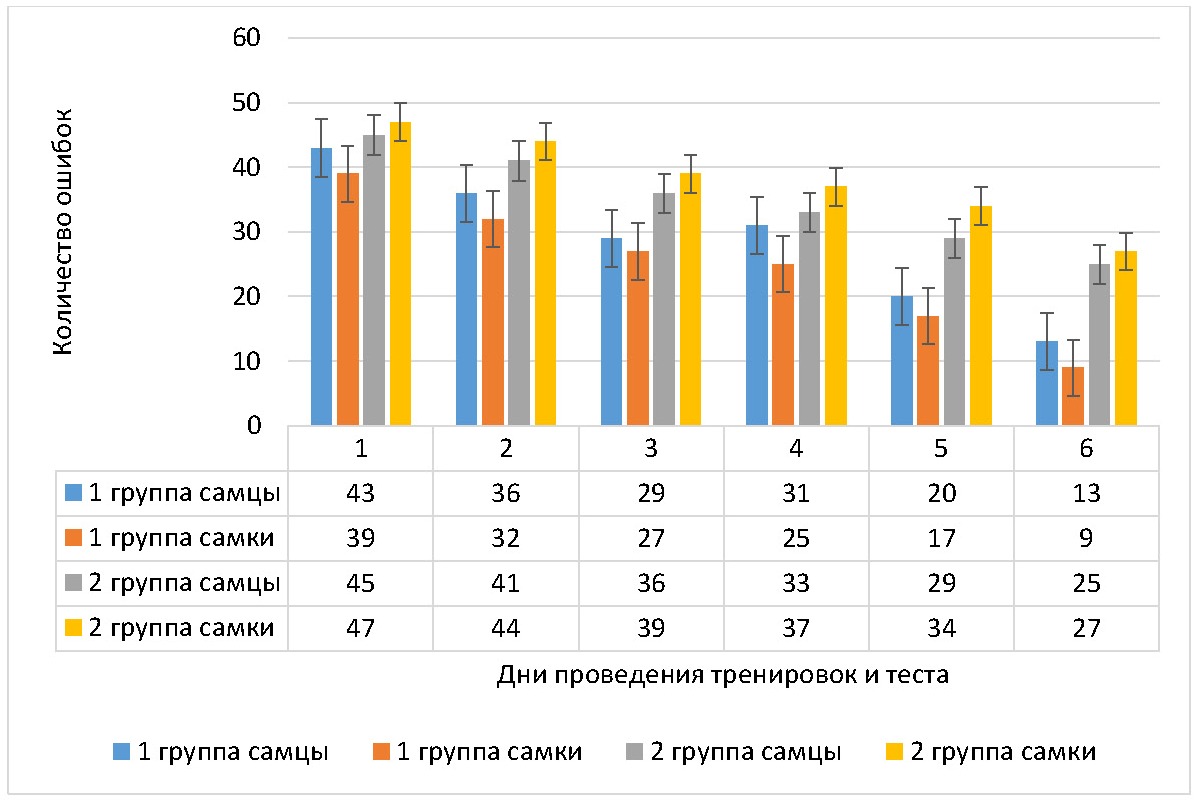
Рисунок 2 - Динамика количества ошибок, совершенных крысами в дни тренировок
Примечание: M±m
У самцов 2 группы количество ошибок на 2 день тренировок снизилось на 8,9% (p <0,034), на 3 день — на 20,0% (p <0,001), на 4 день — на 26,7% (p <0,029), на 5 день — на 35,6% (p <0,001) и на 6 день — на 44,4% (p <0,022) от первоначального значения. У самок 2 группы количество ошибок на 2 день тренировок снизилось на 6,4%, на 3 день — на 17,0% (p <0,001), на 4 день — на 21,9% (p <0,026), на 5 день — на 27,7% (p <0,036) и на 6 день — на 42,6% (p <0,001) от первоначального значения. При этом различия в количестве ошибок с учетом гендерного фактора во 2 группе животных были достоверным в 3 день тренировок (у самок больше ошибок, чем у самцов на 8,3% (p <0,039)), в 4 день тренировок (у самок больше, чем у самцов на 12,1% (p <0,001)), в 5 день теста (у самок больше, чем у самцов на 17,2% (p <0,031).
Применение ДДА (двухфазного дисперсионного анализа) для анализа повторных измерений с учетом межгрупповых (воздействие — ОНГ/контроль; пол – самцы/самки) и внутригрупповых (день обучения — 1–6) факторов выявило значимое влияние фактора ОНГ (F1,243=5,01; p=0,021 (ЛП); F1,243=4,36; p=0,001 для количества ошибок) и сочетания факторов пол и ОНГ (F1,243=1,98; p=0,032 ЛП); F1,243=3,61; p=0,047 для количества ошибок) для ЛП захода в истинную норку и количества совершенных ошибок. Также установлен значимый эффект фактора день обучения на ЛП захода в истинную норку (F4, 139=3,84; p<0,001) и отсутствие значимого взаимодействия факторов ОНГ и день обучения и сочетания факторов ОНГ, пол и день обучения (F1,243=1,75; p>0,75).
Также был выявлен значимый эффект фактора день обучения (F1,243=2,58; p=0,001) и взаимодействие факторов ОНГ и день обучения (F1,243=3,86; p=0,001) для количества ошибок и отсутствие значимого взаимодействия факторов ОНГ, пол и день обучения) (F1,243=0,86; p=0,851). В целом, снижение времени ЛП захода в норку и уменьшение количества ошибок в лабиринте «Барнс» с течением времени свидетельствует о нормальном пространственном обучении животных, но у животных, перенесших ОНГ пространственное обучение запаздывало, по сравнению с контролем.
Результаты теста «Выработка условного пищедобывательного рефлекса на место («Сложный пищевой лабиринт»)» представлены на рисунках 3-4.
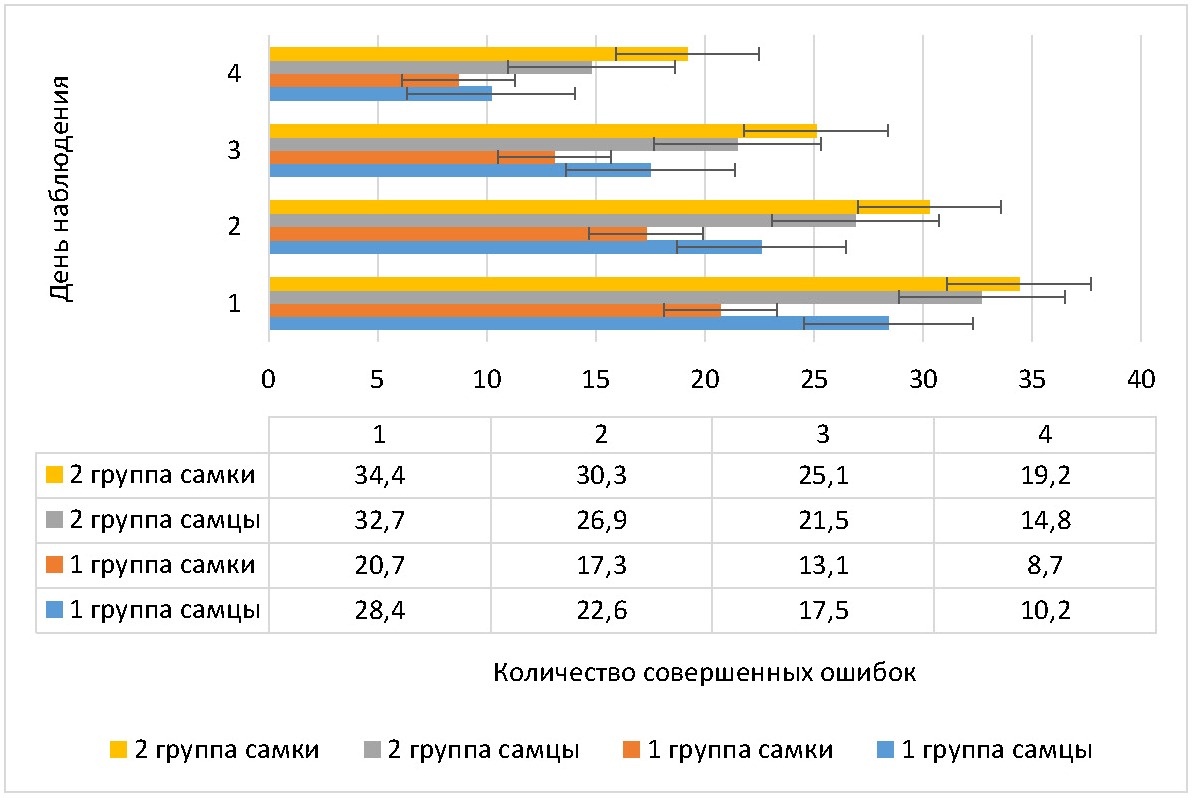
Рисунок 3 - Динамика количества ошибок, совершенных крысами в дни обучения в сложном пищевом лабиринте
Примечание: M±m
У самцов 2 группы количество ошибок на 2 день тренировок снизилось на 17,7% (p <0,001), на 3 день — на 34,3% (p <0,001), на 4 день — на 54,7% (p <0,021) от первоначального значения. У самок 2 группы количество ошибок на 2 день тренировок снизилось на 11,9% (p <0,001), на 3 день — на 27,0% (p <0,001), на 4 день — на 44,2% (p <0,001) от первоначального значения. При этом различия в количестве ошибок с учетом гендерного фактора во 2 группе животных были достоверны на 2 день тренировок (у самок больше ошибок, чем у самцов на 12,6% (p <0,001)), на 3 день тренировок (у самок больше, чем у самцов на 16,7% (p <0,001)), на 4 день тренировок (у самок больше, чем у самцов на 29,7% (p <0,031)).
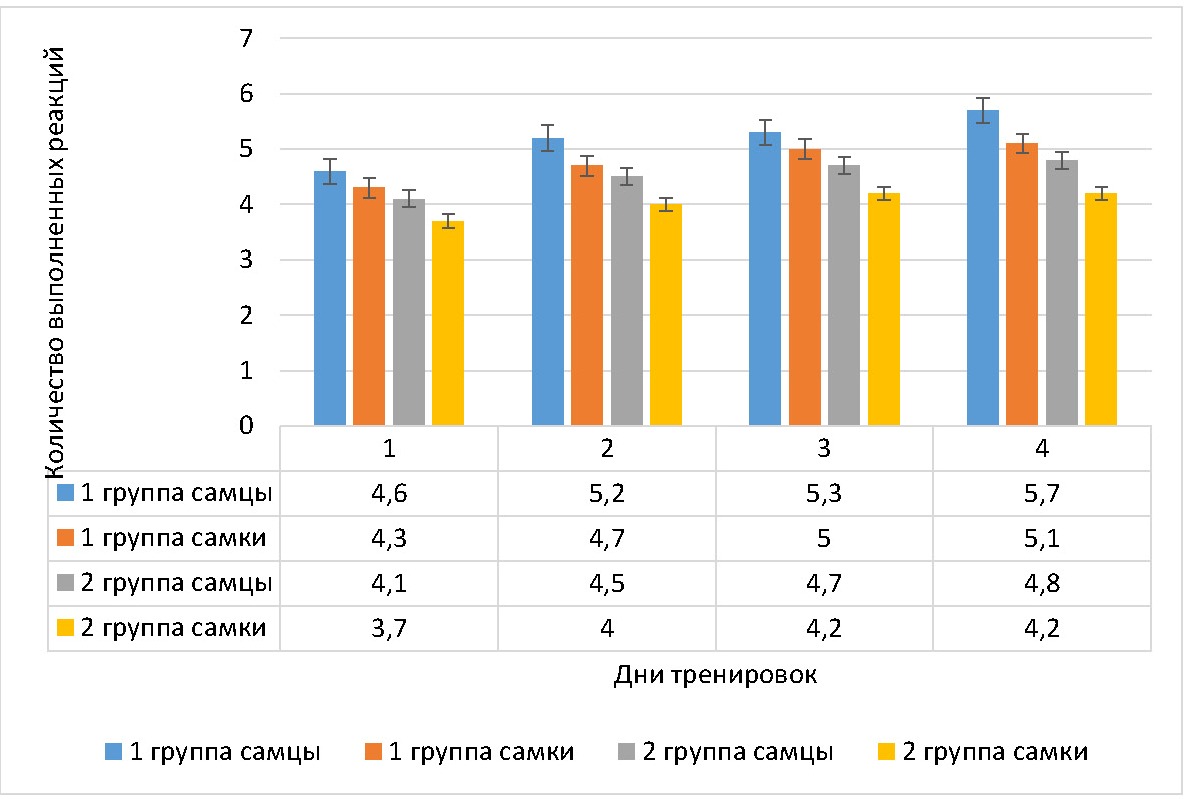
Рисунок 4 - Динамика количества выполненных реакций, совершенных крысами в дни обучения в сложном пищевом лабиринте
Примечание: M±m
У самцов 2 группы количество выполненных реакций на 2 день тренировок увеличилось на 9,8% (p <0,042), на 3 день — на 14,6% (p <0,001), на 4 день — на 17,1% (p <0,021) от первоначального значения. У самок 2 группы количество выполненных реакций на 2 день тренировок увеличилось на 8,1%, на 3 день — на 13,5% (p <0,021), на 4 день — на 13,5% (p <0,001) от первоначального значения. При этом различия в количестве ошибок с учетом гендерного фактора во 2 группе животных были достоверным на 1 день тренировок (у самок меньше количество выполненных реакций, чем у самцов на 9,8% (p <0,001)), на 2 день тренировок (у самок меньше, чем у самцов на 11,1% (p <0,025)), на 3 день тренировок (у самок меньше, чем у самцов на 10,6% (p <0,001)) и на 4 день тренировок (у самок меньше, чем у самцов на 12,5% (p <0,001)).
Анализ поведения животных в сложном пищевом лабиринте проводили с помощью ДДА для повторных измерений с учетом межгрупповых (ОНГ/контроль; самцы/самки) и внутригрупповых (1-4 дни тренировок) факторов. В обеих группах у самок и самцов выявлено значимое влияния фактора день тренировок на количество выполненных реакций (F2, 243=9,5; p <0,001) и количество ошибок (F2, 243=19,8; p <0,002), что свидетельствует об успешном обучении животных. Был отмечен значимый эффект фактора пол на количество выполненных реакций (F1, 243=3,74; p=0,001) и для количества ошибок (F1, 243=5,96; p =0,023). Применение ДДА для повторных измерений показало значимое влияние фактора ОНГ (F1, 243=6,34; p <0,001) и сочетания факторов ОНГ и день обучения (F1, 243=1,78; p <0,001) для количества выполненных реакций и количества ошибок в подгруппе самцов 2 группы. У самок 2 группы с помощью ДДА для повторных измерений установлено значимое влияние фактора ОНГ (F1, 243=8,57; p <0,021) и сочетания факторов ОНГ и день обучения (F1, 243=11,4; p <0,031) для количества ошибок.
4. Обсуждение
По результатам теста «Пространственное обучение в лабиринте «Барнс» на 42-47 сутки жизни было установлено, что:
- Время латентного периода захода в истинные норки животными обоего пола обеих групп снижалось с течением времени эксперимента, но у крыс, подвергшимися ОНГ, на протяжение дней тренировок и теста было достоверно больше, чем у интактных животных, при этом у самок, перенесших гипоксию, было больше, чем у самцов.
- Количество ошибок, совершаемых крысами в дни тренировок и обучения животными обоего пола обеих групп снижалось с течением времени эксперимента, но у крыс, подвергшимися ОНГ, на протяжение дней тренировок и теста было достоверно больше, чем у интактных животных, при этом у самок, перенесших гипоксию, было больше, чем у самцов.
Для оценки способности животных к формированию пространственной памяти проводили анализ изменения стратегий поведения, использованных при поиске убежища. Выделялось три стратегии поведения: случайная (обследование установки случайным образом с пересечением центра лабиринта), последовательная (последовательное движение по краю платформы) и пространственная (с использованием внешних стимулов). И в ходе обучения у интактных животных и крыс, перенесших ОНГ, в целом зафиксировано уменьшение доли случайной стратегии и возрастание долей последовательной и пространственной стратегий, но если у интактных животных это имело линейный характер в зависимости от дня обучения, то у крыс, перенесших ОНГ, особенно самок, изменение стратегий носило хаотичный характер, что свидетельствует о нарушении пространственной памяти, которое в большей степени отразилось на самках, нежели на самцах.
По результатам теста «Сложный пищевой лабиринт» на 42–47 сутки жизни было установлено, что:
- Количество ошибок, совершаемых крысами в дни тренировок и обучения животными обоего пола обеих групп снижалось с течением времени эксперимента, но у крыс, подвергшимися ОНГ, на протяжение дней тренировок и теста было значительно больше ошибок, чем у интактных животных, при этом у самок количество ошибок было больше, чем у самцов.
- Количество выполненных реакций, совершаемых крысами обоего пола обеих групп в дни обучения в сложном пищевом лабиринте с течением времени эксперимента неуклонно росло, но у крыс, подвергшимися ОНГ, на протяжение дней тренировок и теста их было значительно меньше, чем у интактных животных, при этом у самок меньше, чем у самцов.
Полученные результаты свидетельствуют о нарушении выработки пищедобывательного рефлекса на место в сложном лабиринте у крыс, подвергшихся ОНГ, особенно у самок.
Оценка обучения крыс в сложном пищевом лабиринте показала, что ОНГ приводит к нарушению в большей степени у самок крыс способности к выработке пищедобывательного навыка, о чем свидетельствует снижение числа выполненных реакций по сравнению с контролем. В работах Arteni по моделированию гипоксии-ишемии на 7 день жизни крыс, также установлено нарушение рабочей памяти и пространственного обучения на место у самок в возрасте 12 недель в лабиринте Мориса после воздействия
. В работах Morales и соавторов установлено нарушение непространственной рабочей памяти в тесте «Распознавание объекта» после глобальной асфиксии у крыс без указания пола. По данным Buwalda и соавторов установлено нарушение обучения с положительным подкреплением в норковой камере у самцов крыс, перенесших 3 эпизода неонатальной аноксии , самки в данной работе не участвовали.Формирование пространственного обучения приводит к образованию топографического представления об обстановке, основанное на ее очертаниях и распределении стимулов в окружающей среде. Навигационная способность животных помогает сформировать представление об имеющемся самом коротком и эффективном пути через окружающую среду, принимая за основу связи между стимулами. Нарушение способности формировать внутреннее представление об окружающей среде и/или использовать это представление для формирования стратегии навигации может отражаться в неспособности выбрать кратчайший путь к истинной норке в лабиринте Барнса и является показателем нарушений формирования связей в гиппокампе в процессе обучения
. Многие исследования последствий гипоксии у животных и человека указывают на большее влияние гипоксии на самцов, однако имеющиеся результаты не всегда однозначны, а большинство исследовательских групп не учитывают пол животных при изучении неонатальной гипоксии , , . При изучении влияния гипоксии и гипоксии-ишемии на обучение животных, большинство авторов отмечают более выраженные нарушения у самок крыс , . В нашем исследовании в большей степени нарушение выработки навыка в СПЛ, также было отмечено у самок, перенесших ОНГ, хотя у самцов, перенесших ОНГ, тоже наблюдались изменения в обучении и памяти.Согласно клиническим данным, установлены половые различия в частоте встречаемости различных заболеваний, а также в симптомах заболевания и их последствиях у детей, перенесших гипоксию. Экспериментальные исследования также указывают на различия в базальном уровне поведенческой активности у самцов и самок. Половой диморфизм проявляется в процессах обучения и памяти. Так, у самок хуже пространственная память, чем у самцов. Различия в уровне активности и в стресс-опосредованном ответе гипоталамо-гипофизарно-адреналовой оси у самок и самцов зависит от природы стрессорного воздействия. Исследования овариоэктомированных и интактных самок послужили доказательством влияния повышенного уровня эстрогенов в фазу эструса на поведение, нейрохимические маркеры, рост дендритов и ответ на фармакологическое влияние. Половые различия в уровне базальной поведенческой активности, а также сами используемые методики могут оказывать влияние на интерпретацию поведенческих и фармакологических результатов
.Таким образом, наши результаты и литературные данные свидетельствуют о наличии различных по направленности и выраженности поведенческих эффектов в моделях неонатальной гипоксии и пол животных оказывает значимое влияние на проявление определенных поведенческих актов.
5. Заключение
Острая нормобарическая гипоксия приводит к расстройствам поведенческих реакций у крыс адолесцентного возраста, что в тесте «Пространственное обучение в лабиринте «Барнс» выражается дефектами в обучении и памяти, а в тесте «Сложный пищевой лабиринт» — нарушением выработки пищедобывательного рефлекса на место.