КУЛЬТУРА МАЛЬТЫ И ФОРМИРОВАНИЕ ЕВРОПЕОИДНЫХ ПРИЗНАКОВ

Назарова А.Ф.
Доктор биологических наук, старший научный сотрудник, Институт проблем экологии и эволюции РАН, Москва, Россия afnazar@yandex.ru
КУЛЬТУРА МАЛЬТЫ И ФОРМИРОВАНИЕ ЕВРОПЕОИДНЫХ ПРИЗНАКОВ
Аннотация
Вычислена матрица генетических расстояний 11 человеческих популяций Европы, Азии и Америки по 28 аллелям 12 локусов белков, ферментов и групп крови. Построенная по этой матрице дендрограмма показала определенное родство европейских и североазиатских народов, а также американских индейцев. Вычисленная нами матрица генетических расстояний 55 человеческих популяций и построение микроэволюционной дендрограммы этих популяций Европы, Азии, Америки, Африки и Океании, подтвердили это родство. Сопоставление этих данных с общими гаплогруппами митохондриальной ДНК у европейцев, алтайцев и американских индейцев позволило нам предположить о присхождении указанных этносов из одной предковой азиатской палеолитической популяции. Антропологические данные о обнаружении палеолитических костных остатков европеоидов в Сибири подтверждают нашу гипотезу.
Ключевые слова: дифференциация популяций, генетические расстояния, реконструкция, палеолитические популяции, эволюция человеческих популяций.
Nazarova A.F.
Dr. Sci. biol., Senior Scientist
Institute of Ecology and Evolution Problems, Russian Academy of Scienses
Leninsky Prospect, 33, Moscow, 119071, Russia
CULTURE OF MALTA AND FORMING OF CAUCASOID SIGNS
Abstract
The matrix of genetic distances of 28 alleles of 12 loci of proteins, enzymes and blood groups of 11 populations of Europe, Asia and America, and than the matrix of genetic distances of 55 human populations of Europa, Asia, America Africa and Oceania were сalculated. On data of this matrixes we constructed the evolutional dendrogrammes. From this dendrogrammes we suggested that Caucasoids were differentiated with North Mongoloids and Amerinds from Ancient Asiatic population while Middle Palaeolithic in region of Altay and in neighbour regions.The investigations of mitochondrial DNA polymorphism are supported our hypothesis about paleoasiatic origin of North Mongoloids, Caucasoids and Amerinds. The haplogroups of mitochondrial DNA of different human populations of Eurasia and America were marked the way of ancient tribes in their Palaeolithic migrations on map constructed by us. The anthropologic data about discovering of palaeolithic bones of Caucasoids in Siberia are supported our hypothesis.
Key words: population differentiation, genetic distances, reconstruction, Paleolithic populations, human population evolution.
Введение.
Стоянка Мальта времен верхнего палеолита была открыта в 1828 г. в селе Мальта, на берегу р. Белой, левого притока Ангары. Исследование стоянки проводил М.М. Герасимов /Герасимов, 1931, 55-57; Герасимов,1935, 78-123/.
Главным открытием стоянки Мальта является обнаружение погребения палеолитического человека / Герасимов, 1931, 55-57/. Дата фрагмента черепа, полученная в Оксфорде – 19 880 лет назад. Сохранность человеческих костей стоянки Мальта была плохой, поэтому антропологическое изучение костных останков Мальты продолжалось долго. И.И. Гохман обнаружил, что в погребении Мальты было два ребенка, 4-х-летний и годовалый /Алексеев, Гохман, 1991, 54-60/. А.А. Зубов установил, что в морфологии зубов детей из Мальты нет ни одного монголоидного признака (Зубов, Гохман, 2003, 14-23/. И.И. Гохман считал, что захороненные в Мальте дети относились к европеоидной расе.
Известный американский антрополог А. Хрдличка посетил в 1912 г. районы Южной Сибири к западу и востоку от о. Байкал и Монголию. Он считал, что « … в обширных областях Восточной Сибири … имеются многочисленные остатки, входящие теперь как составные элементы в более новые племена или народы. Более древнего (родственного по происхождению может быть с позднейшим палеолитическим европейцем), которое было физически тождественно с американским индейцем и выпустило его из своих недр» /Хрдличка, 1913, 70-75/ .
Материалы и методы.
Мы исследовали полиморфизм белков, ферментов и групп крови популяций алтайцев, эвенков, якутов, чукчей, эскимосов и русских, и вычислили частоты генов исследованных локусов./ Назарова, 2009, 304/. Частоты генов популяций других народов, всего в исследование было взято 55 народов Европы, Азии, Америки, Африки и Океании, мы взяли из литературы. Сводка частот генов всех этих народов приведена нами в / Назарова, Алхутов, 1999, 32. По частотам генов были вычислены генетические расстояния и построены дендрограммы / Nei, 1978, 341-369 ; Назарова, 2009, 304/. Генетическое расстояние –это попытка выразить генетические различия между популяциями по многим признакам одним числом. Сопоставление генетических расстояний и полиморфизма митохондриальной ДНК исследуемых народов Северной Азии / Назарова, 2009, 304/ позволило нам обнаружить определенное генетическое родство ряда европеоидных популяций и популяций Северной Азии (Сибири). Привлечение данных антропологов, обнаруживших костные остатки европеоидов в Сибири в палеолите / Алексеев, 1978,284; Алексеев, Гохман, 1991, 54-60; Бужилова и соавт., 2009, 4-13 /дало нам возможность предположить, что европеоиды дифференцировались от предков северных монголоидов и предков американских индейцев в палеолите в Северной Азии.
Результаты и их обсуждение.
Обнаружение нами европеоидных частот генов белков и ферментов крови /Назарова, Кузнецова,1993, 405-409/ в семи популяциях алтайцев и вычисление генетических расстояний по частотам 28 аллелей белков, ферментов и групп крови 11 человеческих популяций Европы, Азии и Америки: саамов (лопарей), ненцев, нганасан, эвенков, якутов, монголов, алтайцев, русских, финнов, немцев и американских индейцев показало определенную близость европеоидов и северных монголоидов /Назарова,1999, Назарова, 2010, 38-43; Nazarova, 2005, 353-364/. Построенная по матрице генетических расстояний перечисленных выше человеческих популяций Европы, Азии и Америки дендрограмма родства популяций / Назарова,1999, Назарова, 2010, 38-43; Nazarova, 2005, 353-364/ показала, что отделение от общей праазиатской популяции предков американских индейцев произошло раньше других популяций. Предположительно это происходило около 50 тыс. лет назад – это время разделения европеоидов и монголоидов по данным Неи /Неи, 1974,341-369/, вычислившего генетические расстояния крупнейших человеческих рас по большому числу генетических маркеров. Цель нашей работы - показать биологическими методами доказательства формирования европеоидных популяций, северных монголоидов и предков американских индейцев в палеолите в Северной Азии, а также представить небиологические доказательства этого процесса.
После индейцев, по данным дендрограммы (Рис.1) от предковой популяции, обитавшей в регионе Южной Сибири и (или) в соседних регионах Центра Азии, отделились предки саамов. Оставшиеся популяции, указанные на дендрограмме (Рис. 1), далее разделяются на два кластера, в один из которых входят два субкластера: ненцев, нганасан и эвенков, другой – якутов, монголов и алтайцев; во второй кластер входят популяции современных европеоидов – русских, немцев и финнов.
Родство европеоидов и северных монголоидов подтверждается данными о полиморфизме митохондриальной (мт)ДНК: обнаружено наличие одинаковых гаплогрупп у алтайцев и европеоидов Европы – H,J,K,T,U,V,W,F /Деренко и соавт., 2000,1402-1410/, у саамов, эвенков и американских индейцев - гаплогруппа V /Lahermo et al.,1996, 1309-1322/, у алтайцев и американских индейцев - гаплогруппы A, B, C ,D /Сукерник и соавт., 1996,432-439/. Наконец, обнаружение идентичной, и одинаково древней, гаплогруппы X мт ДНК у европеоидов и у американских индейцев /Brown et al.,1998, 1852-1861/ подтвердило общность их происхождения.
Вычисление нами матрицы генетических расстояний по 28 аллелям 12 локусов белков, ферментов и групп крови 55 человеческих популяций, принадлежащих к четырем большим человеческим расам, и обитающим в Европе, Азии, Америке, Африке и Океании /Назарова, Алхутов, 2000, 14-20; Назарова, 2009, 304/, и построение эволюционной дендрограммы по данным этой матрицы (рис. 3) подтвердило наше предположение о возможной дифференциации европеоидных и монголоидных популяций в палеолите в Северной Азии. Раньше всех, как и на малой дендрограмме из (Рис.1), на дендрограмме 55 популяций отделяются предки американских индейцев, затем саамы / Рис. 3/. Нахождение между ветвями индейцев и саамов ветви индийцев видимо, обусловлено тем, что популяция индийцев возникла в результате смешивания европеоидных племен, пришедших с севера в неолите, с имевшими ряд негроидных признаков племенами, населявшими Индостанский полуостров.
В один субкластер с русскими на дендрограмме ( Рис. 3) входят поляки, а в соседний субкластер- иранцы, коми и чуваши, что, возможно связано с тем, что в состав русского этноса вошли около 1000 лет назад племена финно-угорского происхождения – меря, мурома, мещера и другие. В работах акад. Т.И. Алексеевой /Алексеева,1954; Алексеева, 1973, 330 / показано наличие финно-угорского антропологического субстрата в русских популяциях.
Исследование нами митохондриальной ДНК сельской русской популяции Ярославской области выявило наличие практически всех гаплогрупп, присущих русским и даже европеоидам в целом (H, W, I, U,X, T*, T1) /Назарова, Кузнецова, Алхутов,2007, 132-141/.
Предки же немцев мигрировали, по нашему мнению, в Европу из Азии по тому же приблизительно пути, по которому шли уже в историческое время хунну. В работах Сарианиди и соавт./Под. ред. Косарева, Кожина, Дубовой,2004, 469/ описывается фактически открытие поселений древних европеоидов– по мнению /Щербаков, 2002, 364/ – предков германских племен – в Средней Азии, на территории нынешней Туркмении.
В настоящее время исследованы генетические маркеры Y-хромосомы многих человеческих популяций /Klyosov, 2009, 217-256/. Оказалось, что разные этнические популяции обладают четко определенным набором таких маркеров, которые передаются только по мужской линии, поскольку Y-хромосома передается только по мужской линии, от отца к сыну. Эти маркеры являются постоянными, поэтому можно пронаблюдать путь человеческих популяций со времен палеолита до наших дней, а редкое мутирование маркеров позволяет видеть пути разделения (дифференциации) популяций. Далекие предки людей современности около 100 тысяч лет назад вышли из Африки и попали в Азию. Уже в Азии общий маркер Y-хромосомы М 168 изменился на М 89, сейчас этот маркер есть у 90% мужчин - неафриканцев. Дойдя через будущие Иран и Афганистан до гор Памира и Гиндукуша, приобретшие мутацию М 9 около 40 тысяч лет назад предки европейцев и азиатов проникли на Север, в регион Южной Сибири. Там у них возникла мутация Y-хромосомы М 45, около 35 тысяч лет назад. Современным славянским популяциям, например, русским, присуща гаплогруппа Y-хромосомы R 1a1.
По нашему мнению, предки таких славянских популяций, как русские, мигрировали в палеолите из Южной Сибири на Север Сибири, жили некоторое время в Циркумполярной зоне, после чего заселили сначала Восточную, а затем и значительную часть Западной Европы. Недалеко от Красноярска недавно обнаружена лобная кость предположительно, европеоида с датировкой около 28 тыс. лет назад, / Бужилова, Добровольская, Медникова , 2009, 4-13/, а в районе Тобольска – часть стопы неоантропа, датированная 43 тыс. лет назад /24/ Kuzmin, Kosintsev , Razhev , Hodgins, 2009, 91-95/.На стоянке Сунгирь в нынешней Владимирской области были обнаружены европеоиды, жившие там около 25 тысяч лет назад, на стоянке Костенки обнаружены детские европеоидные черепа, датируемые 18 тыс. лет назад. Часть европеоидов–протославян, приобретшая мутацию Y-хромосомы M-173, пошла на Запад, в Европу. В Западной Европе славяне жили до 11-12 веков нашей эры, но с 8-9 веков постепенно вытеснялись с территории будущей Германии на восток. Уже при Карле Великом германские племена вытесняли славянские племена ободритов, полабских славян, поморян, руян с территории будущей Германии на территорию будущей России. О пребывании славян в средние века, и, видимо, ранее, на территории будущей Германии говорится в работах Т.И. Алексеевой /Алексеева, 1973, 330/. И сейчас славянский этнос (лужицкие славяне) обитает на территории Германии.
На рис.2 мы показали возможный путь миграции человеческих популяций, маркированных гаплогруппами митохондриальной ДНК, а также гаплогруппами Y- хромосомы, начиная со среднего палеолита, от места их дифференциации в центре Азии до мест их теперешнего обитания. По нашему мнению, дифференциация этих древних популяций в регионе Южной Сибири могла быть вызвана увеличением числа мутаций по структурным и регуляторным генам в результате глобального катаклизма – взрыва вулкана Тоба в районе нынешней Индонезии около 70 тысяч лет назад, после чего Европа и Северная Америка, и до этого значительно оледеневшие, подверглись чему-то вроде ядерной зимы в результате загрязнения атмосферы. Однако условия жизни в регионе Южной Сибири были лучше, данные палеогеографии говорят о том, что на исходе среднего палеолита оледенение там было связано лишь с вершинами гор, а равнины были свободны от льда /Величко и соавт.,2000, 3-16/.
Миграция древних человеческих популяций могла быть обусловлена изменениями климата, и вызванных этим миграциями животных, на которых охотились люди палеолита. Известно, что предки американского северного оленя (карибу) и предки бизонов в палеолите обитали в регионе Южной Сибири /Верещагин, 1988, 5-32/, и с изменением климата продвинулись на крайний северо-восток Сибири, а затем перешли по существовавшему тогда Берингийскому перешейку в Америку. За ними шли племена предков американских индейцев. Об общей тенденции представителей животного царства к миграциям в древности с Юга Сибири на ее Север говорят данные о
миграциях беспозвоночных (кольчатых червей), и о миграциях птиц, приведенные в книге /Кобельт, 1903/. Есть антропологические данные о существовании европеоидов в палеолите в Северной Азии. Так, академик В.П. Алексеев /Алексеев,1978, 284/ писал о палеолитических черепах с европеоидными признаками в пещере Дунь-Дянь в Китае.
Т.В Томашевич /Томашевич,1988, 119-128/ вычислила градиент распределения частот надглазничных каналов черепа человека (Таблица 1). Оказалось, у саамов, американских индейцев, а также северных монголоидов- якутов, юкагиров, эвенков, чукчей, эскимосов наиболее высокая частота встречаемости надглазничных каналов. У кетов, юкагиров, русских, манси, осетинов, армян частота надглазничных каналов очень близка и варьирует в пределах 30-38 %, это может быть связано с происхождением этих народов из древней палеоазиатской популяции.
По мнению А.Г. Кифишина /Кифишин, 2008, 22-43/, обнаруженная археологами А.П. Окладниковым, В.Е. Ларичевым близ села Суон-Тит на р. Алдан наскальная надпись, сделанная в 18-м тысячелетии до н.э. (культура Мальты) (Рис.4), расшифровывается как протошумерское письмо, появляющееся позже на Бородинской скале под Челябинском на Урале, в протошумерском архиве Каменной Могилы в Приазовье (12-3 тыс. лет до н.э.), и лишь в 3-м тысячелетии до н.э такое письмо появляется в месопотамском Уре. Это подтверждает наши данные о дифференциации европеоидных и монголоидных народов в Азии и их дальнейшей миграции до мест современного обитания, так как видны несколько пунктов многотысячелетней миграции древних народов из региона Сибири через Приазовье в Переднюю Азию.
Заключение.
Таким образом, генетические и антропологические данные, а также данные археолого-лингвистические говорят о том, что европеоиды Европы и Азии, северные монголоиды и американские индейцы происходят из общей праазиатской популяции, обитавшей в палеолите в регионе Алтая, Прибайкалья и в соседних регионах, а в дальнейшем мигрировали до современных мест обитания.
Таблица 1. Распределение частот надглазничных каналов черепа человека
|
Популяции |
Частота надглазничных каналов,% |
Популяции |
Частота надглазничных каналов,% |
|
Поляки |
21,3 (47) |
Русские |
35,9 (204) |
|
Армяне |
30,6 (242) |
Манси |
32,1 (112) |
|
Кеты |
33,3 (38) |
Юкагиры |
38,1 (42) |
|
Осетины |
39,4 (314) |
Башкиры северные |
40,0 (122) |
|
Саамы |
47,4 (221) |
Индейцы |
50,2 (124) |
|
Чукчи береговые |
57,1 (70) |
Эвенки |
57,5 (40) |
|
Эскимосы |
57,4 (302) |
Якуты |
63,0 (144) |
|
Англичане |
17,0 (186) |
Монголы |
48,3 (60) |
|
Словене Новгородской губ.,XII-XIV |
32,5 (152) |
Поляне киевские, IX-XIII в.в. |
25,0 (92) |
|
Этруски, VIII-III в.в. до н.э. |
26,9 (70) |
Голландцы |
21,2 (170) |
|
Древние египтяне |
28,3 |
Негры Руанды |
33,6 (61) |
|
Индийцы Уттар Прадеш |
25,2 (238) |
Негры США |
22,3 (202) |
|
Японцы |
43,2 (1008) |
Корейцы |
46,1 (660) |
|
Австралийцы |
19,0 (122) |
Китайцы |
46,5 (202) |
Подписи к рисункам к статье А.Ф.Назаровой: «Культура Мальты и формирование европеоидных признаков».
Рис. 1. Микроэволюционная дендрограмма 11 человеческих популяций Европы, Азии и Америки. По горизонтальной оси отложены значения генетических расстояний.
1-саамы, 2-ненцы, 3-нганасаны, 4-эвенки, 5-якуты, 6-монголы, 7-алтайцы, 8-русские, 9-финны, 10-немцы, 11-американские индейцы.
Рис.2. Схема палеолитических миграций человеческих популяций, маркированных гаплогруппами митохондриальной ДНК согласно работам разных авторов. В кружках латинскими буквами даны обозначения гаплогрупп мт ДНК.
Рис.3. Микроэволюционная дендрограмма 55 человеческих популяций Европы, Азии, Америки, Африки и Океании. По горизонтальной оси отложены значения генетических расстояний.
Рис.4. Петроглиф у Хана- Шулуун в Прибайкалье, расшифровываемый как палеолитическое протописьмо. Рисунки к статье А.Ф. Назаровой «Культура Мальты и формирование европеоидных признаков».
Рис.1.

1-саамы, ,2-ненцы, 3- нганасаны,4-эвенки,5-якуты,6-монголы,7-алтайцы, 8-русские,9-финны, 10-немцы,11-америнды
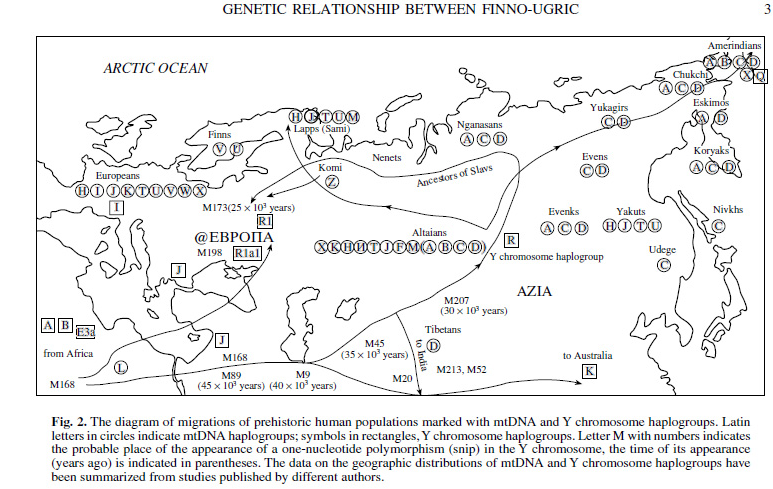
Рис.2.

Рис.3.

Рис. 4.
Литература
1. Алексеев В.П. Палеоантропология земного шара и формирование человеческих рас. М.: Наука, 1978. 284 c.
2. Алексеев В.П., Гохман И.И. Костные остатки детских скелетов из погребения на палеолитической стоянке Мальта. Известия СО АН СССР, сер. истор., филол. и филос., Новосибирск, 1991, вып. 1, с. 54-60..
3. Алексеева Т.И. Антропологический состав Мещеры (к проблеме славяно-финских взаимоотношений в Поволжье). Автореферат канд. дисс. М.:1954.
4. Алексеева Т.И. Этногенез восточных славян по данным антропологии. М.: МГУ. 1973. 330 с.
5. Бужилова А.П., Добровольская М.В., Медникова М.Б. Лобная кость Homo из верхнепалеолитического местонахождения Покровка II в Сибири. Вестник МГУ, сер. Антропология, 2009, №3, стр. 4-13
6. Величко А.А, Кононов Ю.М., Фаустова М.А., 2000. Геохронология, распространение и объем оледенения Земли в последний ледниковый максимум в свете новых данных. Стратиграфия. Геологическая корреляция. 2000, т.8, № 1, 3-16.
7. Верещагин Н.К. Вопросы териологии. Общая и региональная териогеография. М.: Наука, 1988. С. 5-32.
8. Герасимов М.М. Палеолитическая стоянка в Мальте. Сообщения ГАИМК, Л., 1931, вып.11/12, с.55-57.
9. Герасимов М.М. Раскопки палеолитической стоянки в селе Мальте. Палеолит СССР, М.,Л., 1935, с. 78-123.
10. Деренко М.В.,Денисова Г.А., Малярчук Б.А., Дамбуева И.К., Лузина Ф.А., Лотош Е.А., Доржу Ч.М., Карамчакова О.Н., Соловенчук Л.Л., Захаров И.А. Структура генных пулов этнических популяций Алтае-Саянского нагорья на основании данных о поли-морфизме митохондриальной ДНК. Генетика,2001, № 10, 1402-1410.
11. Зубов А.А., Гохман И.И. Некоторые одонтологические данные по верхнепалеолитической стоянке Мальта. Вестник антропологии, 2003, вып. 10, с. 14-23.
12. Кифишин А.Г. Палеоазиатские корни шумеро-вавилонского мифоритуала. В Кн.:Языки и мифоритуалы народов Севера. Новосибирск, Наука, 2008, с. 22-43.
13. Кобельт В. Географическое распространение животных в холодном и умеренном поясах Северного полушария. СПб. 1903.
14. Назарова А.Ф.Популяции, переходные между европеоидами и монголоидами, и возможный путь формирования европеоидов. В кн. Генетический портрет народов мира, М., Липецкое изд-во, 1999, 4-16..
15. Назарова А.Ф. География процессов дифференциации северных монголоидов, европеоидов и предков американских индейцев. В мире научных открытий. 2010. № 2. с.38-43.
16. Назарова А.Ф. Популяционная генетика и происхождение народов Евразии. М., 2009, 304 с.
17. Назарова, А.Ф., Алхутов С.М. Генетические расстояния 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по 28 аллелям 12 локусов белков, ферментов и групп крови. В кн.: Эволюция человеческих популяций, 2000, М., Липецкое издательство, с. 14-20.
18. Назарова А.Ф., Кузнецова М.Г. Генетика популяций алтайцев. Доклады АН, 1993.Т.333.№3.С.405-409.
19. Назарова А.Ф., Кузнецова М.Г., Алхутов С.М. Генетика, экология и демография русской сельской популяции Ярославской области в прошлом и настоящем. В кн.: Социально-модицинские аспекты экологического состояния Центрального экономического района России. Тверь, 2007, с. 132-141.
20. Сукерник Р.И., Шур Т.Г., Стариковская Е.Б. Уоллес Д.С. Изменчивость митохондриальных ДНК у коренных жителей Сибири в связи с реконструкцией эволюционной истории американских индейцев. Генетика, 1996,32, № 3, с.432-439.
21. Томашевич Т.В. Вопр. антропол. 1988 В. 80. С.119-128.
22. У истоков цивилизации. Сборник статей к 75-летию Виктора Ивановича Сарианиди, под ред. Косарева М.Ф., Кожина П.М., Дубовой Н.А., М: изд-во Старый сад, 2004. 469 с.
23. Хрдлычка А. Остатки в Восточной Азии народа, населившего Америку. Труды Троицкосавско-Кяхтинского отделения Приамурского отдела Императорского Русского Географического общества, СПБ, 1913, т. 15, вып. 2, с. 70-75.
24. Щербаков В.И. Асгард- город богов. М. 2001. 364 с.
25. Brown M.D., Hosseini S.H., Torroni A. et al. . mt DNA haplogroup X: an ancient link between Europe/ Western Asia and North America? Amer.J.Hum.Genet., 1998, 63, P.1852-1861.
26. Klyosov A. DNA genealogy, mutation rates, and some historical evidence written in the Y-chromosome: II. Walking the map. J. Genetic Genealogy. 2009, 5, 217-256.
27. Y. V. Kuzmin , P. A. Kosintsev , D. I. Razhev , G. W.L. Hodgins. The oldest directly-dated human remains in Siberia: AMS 14C age of talus bone from the Baigara locality, West Siberian Plain. J. of Humam Evolution, 2009, v.57, 91-95.
28. Lahermo P., Sajantila A., Sistonen P., Lukka M.,Aula P.,Peltonen L.,Savontaus M.-L. The genetic relationship between the Finns and the Finnish Saami (Lapps): analysis of nuclear DNA and mtDNA. Amer.J.Hum.Genet.,1996, 58, P.1309-1322.
29. Nazarova A.F. Genetic Data concerning the Problem of Differentiation of Nothern Mongoloids, American Indians and Caucasoids in the Northern Territory of Eurasia. Anthropologisher Anzeiger, 2005, 63, №4, P.353-364.
30. Nei M. The theory of genetic distance and evolution of human races. Jap.J.Human Genet.1978.V.23. P.341-369.
Список литературы
Алексеев В.П. Палеоантропология земного шара и формирование человеческих рас. М.: Наука, 1978. 284 c.
Алексеев В.П., Гохман И.И. Костные остатки детских скелетов из погребения на палеолитической стоянке Мальта. Известия СО АН СССР, сер. истор., филол. и филос., Новосибирск, 1991, вып. 1, с. 54-60.
Алексеева Т.И. Антропологический состав Мещеры (к проблеме славяно-финских взаимоотношений в Поволжье). Автореферат канд. дисс. М.:1954.
Алексеева Т.И. Этногенез восточных славян по данным антропологии. М.: МГУ. 1973. 330 с.
Бужилова А.П., Добровольская М.В., Медникова М.Б. Лобная кость Homo из верхнепалеолитического местонахождения Покровка II в Сибири. Вестник МГУ, сер. Антропология, 2009, №3, стр. 4-13
Величко А.А, Кононов Ю.М., Фаустова М.А., 2000. Геохронология, распространение и объем оледенения Земли в последний ледниковый максимум в свете новых данных. Стратиграфия. Геологическая корреляция. 2000, т.8, № 1, 3-16.
Верещагин Н.К. Вопросы териологии. Общая и региональная териогеография. М.: Наука, 1988. С. 5-32.
Герасимов М.М. Палеолитическая стоянка в Мальте. Сообщения ГАИМК, Л., 1931, вып.11/12, с.55-57.
Герасимов М.М. Раскопки палеолитической стоянки в селе Мальте. Палеолит СССР, М.,Л., 1935, с. 78-123.
Деренко М.В.,Денисова Г.А., Малярчук Б.А., Дамбуева И.К., Лузина Ф.А., Лотош Е.А., Доржу Ч.М., Карамчакова О.Н., Соловенчук Л.Л., Захаров И.А. Структура генных пулов этнических популяций Алтае-Саянского нагорья на основании данных о поли-морфизме митохондриальной ДНК. Генетика,2001, № 10, 1402-1410.
Зубов А.А., Гохман И.И. Некоторые одонтологические данные по верхнепалеолитической стоянке Мальта. Вестник антропологии, 2003, вып. 10, с. 14-23.
Кифишин А.Г. Палеоазиатские корни шумеро-вавилонского мифоритуала. В Кн.:Языки и мифоритуалы народов Севера. Новосибирск, Наука, 2008, с. 22-43.
Кобельт В. Географическое распространение животных в холодном и умеренном поясах Северного полушария. СПб. 1903.
Назарова А.Ф.Популяции, переходные между европеоидами и монголоидами, и возможный путь формирования европеоидов. В кн. Генетический портрет народов мира, М., Липецкое изд-во, 1999, 4-16.
Назарова А.Ф. География процессов дифференциации северных монголоидов, европеоидов и предков американских индейцев. В мире научных открытий. 2010. № 2. с.38-43.
Назарова А.Ф. Популяционная генетика и происхождение народов Евразии. М., 2009, 304 с.
Назарова, А.Ф., Алхутов С.М. Генетические расстояния 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по 28 аллелям 12 локусов белков, ферментов и групп крови. В кн.: Эволюция человеческих популяций, 2000, М., Липецкое издательство, с. 14-20.
Назарова А.Ф., Кузнецова М.Г. Генетика популяций алтайцев. Доклады АН, 1993.Т.333.№3.С.405-409.
Назарова А.Ф., Кузнецова М.Г., Алхутов С.М. Генетика, экология и демография русской сельской популяции Ярославской области в прошлом и настоящем. В кн.: Социально-модицинские аспекты экологического состояния Центрального экономического района России. Тверь, 2007, с. 132-141.
Сукерник Р.И., Шур Т.Г., Стариковская Е.Б. Уоллес Д.С. Изменчивость митохондриальных ДНК у коренных жителей Сибири в связи с реконструкцией эволюционной истории американских индейцев. Генетика, 1996,32, № 3, с.432-439.
Томашевич Т.В. Вопр. антропол. 1988 В. 80. С.119-128.
У истоков цивилизации. Сборник статей к 75-летию Виктора Ивановича Сарианиди, под ред. Косарева М.Ф., Кожина П.М., Дубовой Н.А., М: изд-во Старый сад, 2004. 469 с.
Хрдлычка А. Остатки в Восточной Азии народа, населившего Америку. Труды Троицкосавско-Кяхтинского отделения Приамурского отдела Императорского Русского Географического общества, СПБ, 1913, т. 15, вып. 2, с. 70-75.
Щербаков В.И. Асгард- город богов. М. 2001. 364 с.
Brown M.D., Hosseini S.H., Torroni A. et al. . mt DNA haplogroup X: an ancient link between Europe/ Western Asia and North America? Amer.J.Hum.Genet., 1998, 63, P.1852-1861.
Klyosov A. DNA genealogy, mutation rates, and some historical evidence written in the Y-chromosome: II. Walking the map. J. Genetic Genealogy. 2009, 5, 217-256.
Y. V. Kuzmin , P. A. Kosintsev , D. I. Razhev , G. W.L. Hodgins. The oldest directly-dated human remains in Siberia: AMS 14C age of talus bone from the Baigara locality, West Siberian Plain. J. of Humam Evolution, 2009, v.57, 91-95.
Lahermo P., Sajantila A., Sistonen P., Lukka M.,Aula P.,Peltonen L.,Savontaus M.-L. The genetic relationship between the Finns and the Finnish Saami (Lapps): analysis of nuclear DNA and mtDNA. Amer.J.Hum.Genet.,1996, 58, P.1309-1322.
Nazarova A.F. Genetic Data concerning the Problem of Differentiation of Nothern Mongoloids, American Indians and Caucasoids in the Northern Territory of Eurasia. Anthropologisher Anzeiger, 2005, 63, №4, P.353-364.
Nei M. The theory of genetic distance and evolution of human races. Jap.J.Human Genet.1978.V.23. P.341-369.
Список литературы
Алексеев В.П. Палеоантропология земного шара и формирование человеческих рас. М.: Наука, 1978. 284 c.
Алексеев В.П., Гохман И.И. Костные остатки детских скелетов из погребения на палеолитической стоянке Мальта. Известия СО АН СССР, сер. истор., филол. и филос., Новосибирск, 1991, вып. 1, с. 54-60.
Алексеева Т.И. Антропологический состав Мещеры (к проблеме славяно-финских взаимоотношений в Поволжье). Автореферат канд. дисс. М.:1954.
Алексеева Т.И. Этногенез восточных славян по данным антропологии. М.: МГУ. 1973. 330 с.
Бужилова А.П., Добровольская М.В., Медникова М.Б. Лобная кость Homo из верхнепалеолитического местонахождения Покровка II в Сибири. Вестник МГУ, сер. Антропология, 2009, №3, стр. 4-13
Величко А.А, Кононов Ю.М., Фаустова М.А., 2000. Геохронология, распространение и объем оледенения Земли в последний ледниковый максимум в свете новых данных. Стратиграфия. Геологическая корреляция. 2000, т.8, № 1, 3-16.
Верещагин Н.К. Вопросы териологии. Общая и региональная териогеография. М.: Наука, 1988. С. 5-32.
Герасимов М.М. Палеолитическая стоянка в Мальте. Сообщения ГАИМК, Л., 1931, вып.11/12, с.55-57.
Герасимов М.М. Раскопки палеолитической стоянки в селе Мальте. Палеолит СССР, М.,Л., 1935, с. 78-123.
Деренко М.В.,Денисова Г.А., Малярчук Б.А., Дамбуева И.К., Лузина Ф.А., Лотош Е.А., Доржу Ч.М., Карамчакова О.Н., Соловенчук Л.Л., Захаров И.А. Структура генных пулов этнических популяций Алтае-Саянского нагорья на основании данных о поли-морфизме митохондриальной ДНК. Генетика,2001, № 10, 1402-1410.
Зубов А.А., Гохман И.И. Некоторые одонтологические данные по верхнепалеолитической стоянке Мальта. Вестник антропологии, 2003, вып. 10, с. 14-23.
Кифишин А.Г. Палеоазиатские корни шумеро-вавилонского мифоритуала. В Кн.:Языки и мифоритуалы народов Севера. Новосибирск, Наука, 2008, с. 22-43.
Кобельт В. Географическое распространение животных в холодном и умеренном поясах Северного полушария. СПб. 1903.
Назарова А.Ф.Популяции, переходные между европеоидами и монголоидами, и возможный путь формирования европеоидов. В кн. Генетический портрет народов мира, М., Липецкое изд-во, 1999, 4-16.
Назарова А.Ф. География процессов дифференциации северных монголоидов, европеоидов и предков американских индейцев. В мире научных открытий. 2010. № 2. с.38-43.
Назарова А.Ф. Популяционная генетика и происхождение народов Евразии. М., 2009, 304 с.
Назарова, А.Ф., Алхутов С.М. Генетические расстояния 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по 28 аллелям 12 локусов белков, ферментов и групп крови. В кн.: Эволюция человеческих популяций, 2000, М., Липецкое издательство, с. 14-20.
Назарова А.Ф., Кузнецова М.Г. Генетика популяций алтайцев. Доклады АН, 1993.Т.333.№3.С.405-409.
Назарова А.Ф., Кузнецова М.Г., Алхутов С.М. Генетика, экология и демография русской сельской популяции Ярославской области в прошлом и настоящем. В кн.: Социально-модицинские аспекты экологического состояния Центрального экономического района России. Тверь, 2007, с. 132-141.
Сукерник Р.И., Шур Т.Г., Стариковская Е.Б. Уоллес Д.С. Изменчивость митохондриальных ДНК у коренных жителей Сибири в связи с реконструкцией эволюционной истории американских индейцев. Генетика, 1996,32, № 3, с.432-439.
Томашевич Т.В. Вопр. антропол. 1988 В. 80. С.119-128.
У истоков цивилизации. Сборник статей к 75-летию Виктора Ивановича Сарианиди, под ред. Косарева М.Ф., Кожина П.М., Дубовой Н.А., М: изд-во Старый сад, 2004. 469 с.
Хрдлычка А. Остатки в Восточной Азии народа, населившего Америку. Труды Троицкосавско-Кяхтинского отделения Приамурского отдела Императорского Русского Географического общества, СПБ, 1913, т. 15, вып. 2, с. 70-75.
Щербаков В.И. Асгард- город богов. М. 2001. 364 с.
Brown M.D., Hosseini S.H., Torroni A. et al. . mt DNA haplogroup X: an ancient link between Europe/ Western Asia and North America? Amer.J.Hum.Genet., 1998, 63, P.1852-1861.
Klyosov A. DNA genealogy, mutation rates, and some historical evidence written in the Y-chromosome: II. Walking the map. J. Genetic Genealogy. 2009, 5, 217-256.
Y. V. Kuzmin , P. A. Kosintsev , D. I. Razhev , G. W.L. Hodgins. The oldest directly-dated human remains in Siberia: AMS 14C age of talus bone from the Baigara locality, West Siberian Plain. J. of Humam Evolution, 2009, v.57, 91-95.
Lahermo P., Sajantila A., Sistonen P., Lukka M.,Aula P.,Peltonen L.,Savontaus M.-L. The genetic relationship between the Finns and the Finnish Saami (Lapps): analysis of nuclear DNA and mtDNA. Amer.J.Hum.Genet.,1996, 58, P.1309-1322.
Nazarova A.F. Genetic Data concerning the Problem of Differentiation of Nothern Mongoloids, American Indians and Caucasoids in the Northern Territory of Eurasia. Anthropologisher Anzeiger, 2005, 63, №4, P.353-364.
Nei M. The theory of genetic distance and evolution of human races. Jap.J.Human Genet.1978.V.23. P.341-369.