Листовая пластина Липы мелколистной: низкий фитнес, малая величина и высокая асимметрия
Листовая пластина Липы мелколистной: низкий фитнес, малая величина и высокая асимметрия
Аннотация
Многие исследователи предполагают, что флуктуирующая асимметрия (ФА) – это мера онтогенетического шума, который является интегральной величиной биохимических и физиологических факторов, дестабилизирующих гомеостаз организма. Предложена регрессионная модель интегрального индекса ФА популяций центральной и периферической частей ареала липы мелколистной (Tilia cordata Mill), которая включает такие факторы как: а) эксцесс распределения коэффициентов корреляции пластических признаков (FS), б) коэффициент вариации размеров этих признаков (CV), в) шум пластической изменчивости, как долю вариации признаков без флуктуирующей асимметрии (DN). Наиболее высокий коэффициент регрессии имел эксцесс (b=0.3), далее – CV (b=0.21) и DN (b=0.20; везде p<<0.05). CV и FS были выше в северо-западном регионе по сравнению с московской областью (p<<0.05). Популяции не отличались по значению ФА, отнесенному к коэффициенту вариации признаков. В северо-западном регионе отмечен больший шум развития на фоне низкого показателя фитнеса, чем в Подмосковье (p=0,0001). Индекс ФА был существенно выше в северо-западном регионе из-за малой площади листьев (отрицательная корреляция, p<0.05) и мог отражать генетический компонент флуктуационной изменчивости. Таким образом, для пластических признаков продемонстрирована связь: низкий фитнес – малая величина – высокая асимметричность. Авторы предполагают, что, асимметрия формы, в отличие от асимметрии мерных признаков, имеет скорее средовое происхождение, тогда как асимметрия метрических признаков зависит от морфогенов, реализующихся под действием факторов среды.
1. Введение
Периферийные и центральные популяции древесных растений показывают пример фенотипической пластичности, например, холодный климат приводит к мелким размерам и высокой изменчивости формы листовых пластин. Онтогенетический шум, связан с флуктуационной изменчивостью, имеет, по крайней мере, две причинные основы: детерминистскую и стохастическую или осциляторную, которые могут иметь как генетическое, так и фенотипическое происхождение . Возможно, генетический шум развития блокирует нежелательные пульсации ФА. В таких случаях не происходит ожидаемого эффекта увеличения ФА при повышении индустриального или иного стресса. Альтернативные или дополнительные точки зрения объясняются растянутостью во времени ответной реакции . Таким образом, генетический шум развития как буфер может нейтрализовать нежелательные биохимические процессы, приводящие к разбалансировке морфогенеза билатеральных структур.
Доля онтогенетического шума может быть разной в зависимости от выбранных признаков, таксономического положения вида и его эволюционного креода развития.
Предыдущие работы показали, что популяционная изменчивость липы мелколистной, например, флуктуирующей асимметрии (нормирующая разность мерных признаков) была выше в северных широтах (Кольский полуостров) по сравнению со средней полосой России на фоне значительно меньших размеров листьев в северных популяциях. Методом геометрической морфометрии, основанным на множестве меток по краям листовой пластины, выявлена более высокая флуктуирующая асимметрия в популяциях средней полосы , . Учитывая эти результаты, мы посчитали важным обратить внимание на определение жизнеспособности популяций липы.
В литературе сложилось противоречивое мнение о связи фитнес – стабильность развития. Существует несколько мнений относительно природы и проявления фитнеса растительного организма. Чаще это способность к полноценной генерации потомства и его эволюции. Мы использовали парные коэффициенты корреляции метрических признаков, как условный показатель фитнеса растения на уровне конкретного вегетативного органа – листовой пластины. Дополнительно включали выборочную характеристику – эксцесс распределения.
Шум развития, вероятно, является отражением шума пластической изменчивости, расчитываемой на основе вариации метрических признаков. В данной статье предлагается определение онтогенетического шума, как доли пластической изменчивости, лишенной флуктуирующей асимметрии . Методом множественной регрессии оценивалось влияние эксцесса распределения корреляционных коэффициентов, шума развития и вариации признаков на величину ФА.
Листовая пластина липы мелколистной удобна для исследования, так как обладает достаточно большим числом (6) билатерально-симметричных признаков. Места сбора листьев находились как в индустриальных локалитетах, так и в биотопах свободных от антропогенного стресса.
Интерес вызывает генетический шум развития, который может проявляться отклонением в ФА на периферии ареалов . Таким образом, в задачу настоящей работы входило определение факторов, оказывающих наибольшее влияние на флуктуирующую асимметрию в популяциях разных частей ареала липы мелколистной, а также сравнение в них шума развития как пластической изменчивости с учетом фитнеса растения.
2. Методы и принципы исследования
В работе использовались гербарные материалы из популяций, находящихся в разных физико-географических условиях: г. Мурманск (68°58′00′′ с. ш., 33°05′00″ в. д.), г. Апатиты (67°34′03″ с. ш., 33°23′36″ в. д.), г. Петрозаводск (61°47′46′′ с. ш., 34°20′57′′ в. д.), г. Санкт-Петербург (59°56′19″ с. ш., 30°18′50″ в. д.), г. Москва (Измайловский район, 55°45′07″ с. ш., 37°36′56″ в. д.) и г. Орехово-Зуево (55°48′24″ с. ш., 38°57′43″ в. д.) и г. Можайск (55°30′06″ с. ш., 36°01′37″ в. д.) Московской области. Предыдущие исследования показали ответную реакцию на загрязнения только по некоторым признакам, таким как расстояние, между основаниями первой и второй жилок первого порядка и ширина половины листа, поэтому, важно было оценить популяции по интегрированному показателю. В указанных локалитетах листья липы мелколистной собирались в августе-сентябре 2014-2017 годов с нижних частей крон, равномерно со всех сторон по 10 листовых пластин с каждого из десяти деревьев.
После сканирования (300 dpi) изображения листьев двукратно измерялись с помощью экранного дигитайзера TPSdig2 (Rholf, 2017). Результаты измерений 5-6 мерных признаков сначала заносились в таблицы Excel, затем – в таблицы STATISTICA 10 (Statsoft Ink.), где выполнялись основные анализы описательной статистики. Проверка на нормальность проводилась с помощью критерия Колмогорова-Смирнова (тест К-С). Коэффициент корреляции между значениями величины признака и ФА2 (FA2=|L–R|/(L+R)) находился по формуле Пирсона для нормально распределенных выборок или по формуле Спирмена для выборок, отклонявшихся от нормального распределения.
Показатель фитнеса определялся в виде среднего арифметического взвешенного исходя из корреляционной матрицы для всех признаков. Учитывались лишь статистически значимые коэффициенты, среднее значение которых относилось к общему числу корреляционных пар.
Признаки с направленной асимметрией (НА) исключались из работы (t тест, H0: L=R; 20% случаев из 27 популяций). Присутствия антисимметрии (отрицательный коэффициент корреляции между L и R) получено не было.
Для определения шума развития мы использовали долю вариации признаков, лишенную ФА и отнесенную к размеру признака:
DN=(1–FA/CV)/(L+R)/2,
где:
DN – шум пластической изменчивости;
FA – индекс ФА2;
CV – коэффициент вариации величины признака;
(L+R)/2 – средний размер признака.
Нормализация и генерирование 100 значений проводились в Excel с учетом среднего значения и стандартного отклонения. Генерировались три выборки по 100 значений для каждого года сбора с 2014 по 2017:
1) коэффициент вариации размеров признаков (CV, для удобства отнесенный к 100),
2) показатель фитнеса (FS) в виде эксцесса распределения,
3) показатель пластического шума развития (DN).
С учетом расположения популяций (2 региона) и года сбора (4 года) общая таблица включала 800 строк. Категории «год» и «место сбора» были кодовыми переменными; FA, CV, FS, DN служили числовыми переменными. Уровень значимости нулевой гипотезы принимался за 95%.
3. Результаты
3.1. Описательная статистика
Листовые пластины липы мелколистной, как отмечалось, варьируют по величине, особенно в северных районах, в зависимости от высоты над уровнем моря. Тест Левина на гомогенность дисперсии показал различие между популяциями северо-западного и московского регионов (р<<0.000), при статистически значимом различии в средних (р<0.000) . Значения ФА2, полученные по нормализующей формуле, обладали различными дисперсиями, характеризовавшими выборки. После генерации нормальных выборок, проверка тестом Колмогорова показала нормальное распределение в ФА, кроме показателей CV и FS (московский регион; p<0.05; табл. 1).
Таблица 1 - Описательная статистика
Место/индекс | Среднее | Стандартная ошибка | Дисперсия | Эксцесс | Скос | K-S тест, p |
Московский регион, n = 400 | ||||||
FA | 0,058 | 0,00 | 0,00 | 0,03 | 0,75 | p>0,20 |
CV | 0,155 | 0,00 | 0,00 | 0,05 | 1,53 | p<0,010 |
FS | 0,245 | 0,01 | 0,03 | 0,38 | 0,41 | p<0,05 |
DN | -0,121 | 0,05 | 0,82 | – 0,11 | –0,36 | p>0,20 |
Северо-западный регион, n = 400 | ||||||
FA | 0,08 | 0,00 | 0,00 | – 0,13 | 0,54 | p>0,20 |
CV | 0,23 | 0,00 | 0,01 | 0,01 | 0,06 | p>0,20 |
FS | 0,21 | 0,00 | 0,01 | 0,12 | 0,0 | p>0,20 |
DN | 0,92 | 0,05 | 1,14 | 0,04 | 0,33 | p>0,20 |
Примечание: FA – флуктуирующая асимметрия (ФА); CV – коэффициент вариации величин признаков; FS – показатель фитнеса; DN – показатель шума развития
3.2. Флуктуирующая асимметрия и коэффициент вариации признаков
ФА (F=1871.4; р<0,001) и коэффициент вариации величины признаков (F=238.4; р<0,001) преобладали в популяциях северо-западной части России по сравнению с подмосковным регионом. ФА признаков отрицательно коррелировала с их величиной. Листья в северо-западном регионе были меньше по размеру и обладали более высокой ФА (рис. 1).
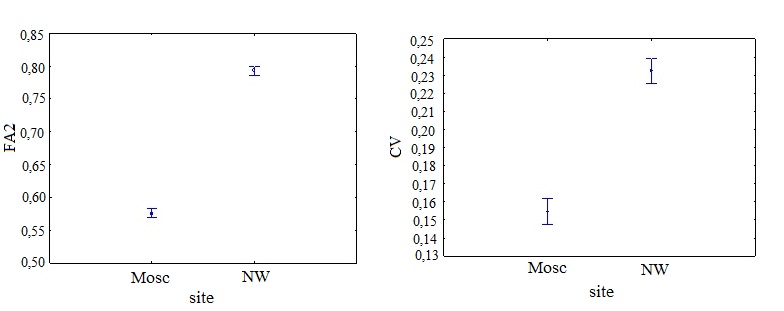
Рисунок 1 - Зависимость ФА (FA2) и вариации признаков (CV) от расположения популяций (site)
Примечание: ФА признаков отрицательно коррелировала с их величиной; листья в Северо-западном регионе были меньше и обладали высокой ФА; Mosc – Московский регион; NW – Северо-западный регион
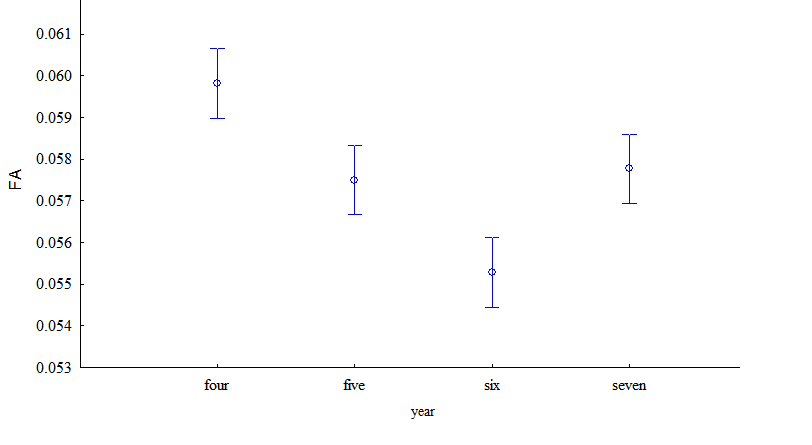
Рисунок 2 - Зависимость ФА (FA) и вариации признаков (CV) от года сбора материала (2014-2017)
3.3. Коррелятивные особенности
Получены корреляционные коэффициенты по всем годам исследования всех популяций (табл.2). Установлена положительная корреляция между ФА и коэффициентом вариации величин признаков, шум развития отрицательно коррелировал с ФА и сильно положительно – с CV.
Таблица 2 - Коэффициенты корреляции
r Спирмена | r Пирсона | |||||||||
Московская область | ||||||||||
FA | CV | FS | DN | FA | CV | FS | DN | |||
FA | х | FA | х | |||||||
CV | – 0,15* | х | CV | –0,14* | х | |||||
FS | 0,1* | – 0,1 | х | FS | 0,09 | –0,08 | х | |||
DN | –0,53* | 0,20* | – 0,14* | х | DN | –0,38* | 0,84* | -0,1* | х | |
Северо-западный регион | ||||||||||
FA | х | FA | х | |||||||
CV | –0,02 | х | CV | –0,00 | х | |||||
FS | 0,01 | 0,02 | х | FS | –0,02 | –0,00 | х | |||
DN | –0,23* | 0,92* | 0,03 | х | DN | –0,1 | 0,44* | 0,05 | х | |
Примечание: FA – индекс флуктуирующей асимметрии, CV – коэффициент вариации, FS – эксцесс вариации фитнеса, DN – индекс шума развития
Индекс фитнеса был выше в Московской области (F=14.7; р<0,001). Он слабо коррелировал (коэффициент Пирсона r=0.1) с FA2, что на наш взгляд говорит о специфичности этого показателя и необходимости его дальнейшего изучения. Отмечено отсутствие связи «фитнес–шум развития» в северо-западном регионе.
3.4. Зависимость шума развития от года сбора листовых пластин и географического расположения популяций
Величина шума развития зависела от расположения популяций (F=321.9; р<0,001) и года сбора листьев (F=9.2; р=0,00001). Наибольшие значения получены в 2015 и 2016 годах (рис. 3).
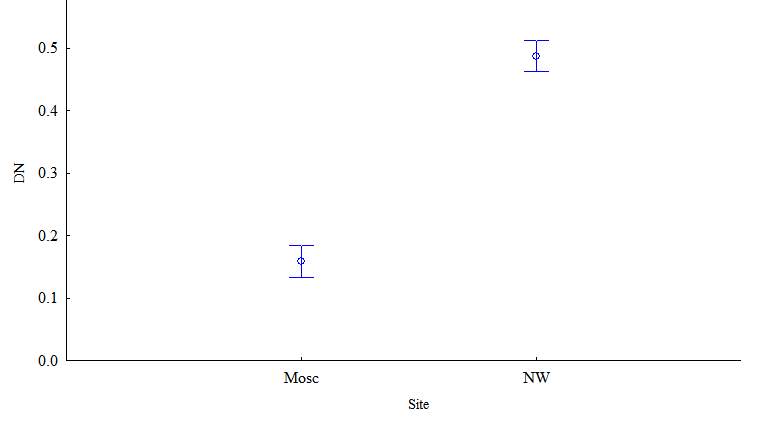
Рисунок 3 - Зависимость шума развития от географического расположения популяций
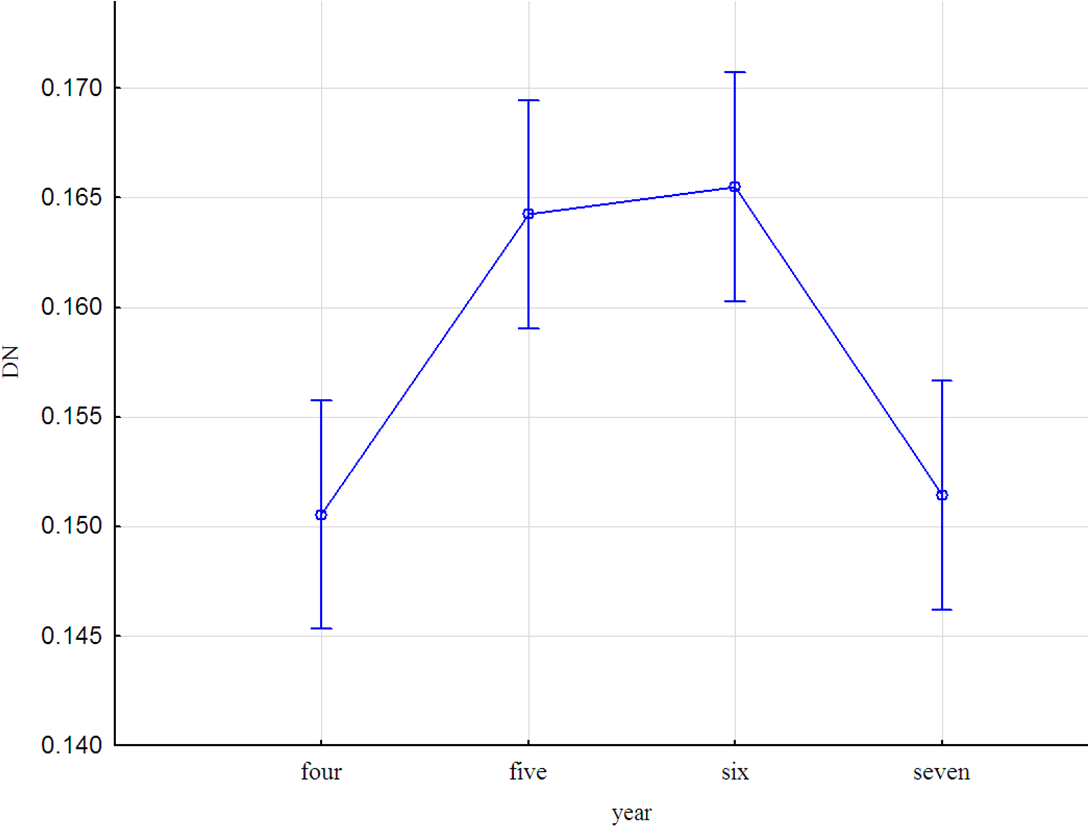
Рисунок 4 - Зависимость шума развития от года сбора листовых пластин
3.5. Многофакторный регрессионный анализ
Чтобы определить, от чего зависит флуктуирующая асимметрия, кроме географического расположения популяций и года сбора, был поставлен регрессионный анализ с использованием всех переменных (табл. 3). Получены коэффициенты множественной регрессии: наиболее значимым был фактор «фитнес» (b=0.3), затем – коэффициенты вариации размера признаков (b=0.21) и шум развития (b=0.20; р<<0,001; константа b≠0).
Таблица 3 - Факторы регрессии, объясняющие вариацию флуктуирующей асимметрии
Индексы | SS | Degr, of | MS | F | p |
DN | 0,003 | 1 | 0,003 | 26,477 | 0,000 |
FS | 0,011 | 1 | 0,011 | 89,746 | 0,000 |
CV | 0,004 | 1 | 0,004 | 33,521 | 0,000 |
Error | 0,096 | 795 | 0,000 |
|
|
Коэффициент детерминации был равен R2=0,28±0.01, что говорит о 28% объясненной дисперсии вариации ФА. Наибольший эффект играла вариация эксцесса распределения величин признаков (F=89,8), то есть не сама величина корреляционной связи между признаками, а форма распределения случайной величины. Шум развития, влиял различно на популяции. В московском регионе он имел значительно больший эффект (F=122,8; p<<0.001), чем в северо-западных районах России (F=4.2; p≤0.05).
3.6. Выводы
1. Северо-запад страны – краевая часть ареала липы мелколистной, включающая ее изолированные популяции за полярным кругом. Адаптация здесь связана с уменьшением размеров листовых пластин и увеличением изменчивости признаков, вариативности формы и увеличением индекса ФА.
2. Вариация, как изменчивость признаков, была тесно связана с особенностями года. Метрические признаки листовых пластин не коррелировали с индексом ФА, момент распределения случайной величины (эксцесса) оказывал существенное влияние на отклик ФА в регрессионной модели.
3. Величина шума развития, как доли пластической изменчивости, была более высокой в северо-западном регионе и могла обладать не только средовой, но генетической вариационной составляющей.
4. Обсуждение
В нашей предыдущей работе показана отрицательная связь между двумя видами изменчивости липы широколистной. Обратная корреляция ФА – пластическая изменчивость продемонстрирована на примере лишь одного билатерально симметричного признака, в настоящей работе эта же тенденция подтверждена для интегрированного показателя липы мелколистной. По нашему мнению, скорее пластическая изменчивость оказывала влияние на ФА по схеме: климатический стресс (холодный год) – пластическая изменчивость (уменьшение размера листовых пластин) – повышение ФА. Полученная негативная корреляционная связь между ФА и пластической изменчивостью скорее говорит в пользу возможности локализация генов в одном генетическом локусе у представителей рода Tilia L. Подобная реакция на климатический стресс в виде увеличения ФА характерна для древесных растений и других семейств.
Мы согласны с авторами в том, что не только коррелятивные особенности между признаками, но и характер распределения случайной величины, может помочь понять происхождение шума развития. По нашему мнению, эксцесс, как и другие характеристики распределения, могут служить для изучения стабильности развития и онтогенетического шума.
Существует две точки зрения на связь пластической и флуктуационной изменчивости: независимое и взаимозависимое их проявление . У растений, ведущих существование, фиксированное к субстрату, освещение и температура – важнейшие экофакторы, индуцирующие и моделирующие как детерминистские, так и стохастические процессы шума онтогенетического развития. Для онтогенетического шума, который, как предполагается, призван нейтрализовать возмущения в биологической системе, скорее характерна высокая вариативность. Ответ на стресс и механизм буферной нейтрализации стресса протекает динамически и их затруднительно разделить . Флуктуирующая асимметрия – лишь случайный сигнал, отражающий индуцированный всплеск шума развития.
Экологами и биостатистиками предпринимаются попытки определить шум развития исходя из свойства пластической изменчивости. Моделирование широко применяется для изучения роли пластической изменчивости в эволюции, а также для определения факторов, лимитирующих адаптивную фенотипическую пластичность . Сторонниками корреляции пластичности и нестабильности смоделировано влияние на наследственность и фитнес . На основе молекулярно-генетической базы данных как независимых, так и плейотропно зависимых локусов генов в различных условиях среды, и сделан вывод о негативной корреляции между ФА и пластической изменчивостью в случае генетической независимости обоих генов. Липа мелколистная имеет в геноме 82 хромосомы, что по сравнению, например, с другими семействами (Березовые – 18) увеличивает вероятность индивидуального генетического контроля пластической и флуктуационной изменчивости. Согласно модели, предложенной Scheiner, если пластичность и нестабильность определялись одними и теми же локусами, то нестабильность служит ограничением для пластической изменчивости , такое предположение также заслуживает внимания и изучения.
5. Заключение
В северо-западных популяциях присутствовала направленная асимметрия и высокая изменчивость формы, в отличие от центральной части ареала липы мелколистной. В задачу последующих исследований будет входить изучение генетически близких популяций для понимания роли негенетического компонента шума развития. Метод геометрической морфометрии не показал существенного различия во флуктуирующей асимметрии формы. Мы полагаем, что асимметрия формы, не совпадающая с асимметрией признаков, также может нести информацию о фенотипическом компоненте шума развития.