К ВОПРОСУ О ДЕМОНСТРАТИВНЫХ ВОКАЛИЗАЦИЯХ БОЛЬШОЙ ВЫПИ (BOTAURUS STELLARIS)

Яновский И.Ю.
Аспирант, Санкт-Петербургский Государственный университет
К ВОПРОСУ О ДЕМОНСТРАТИВНЫХ ВОКАЛИЗАЦИЯХ БОЛЬШОЙ ВЫПИ (BOTAURUS STELLARIS)
Аннотация
Работа посвящена изучению вариабельности частотно-временных характеристик демонстративных вокализаций большой выпи (Botaurus stellaris), представлен краткий ретроспективный обзор основных работ по данной тематике.
Демонстративная вокализация самцов большой выпи рассматривается, как песня, функционально аналогичная песне воробьиных.
Исследования проводились на территории Псковского Поозерья в период активной вокализации самцов. По результатам наблюдений приведены значения параметрических измерений (которые могут быть использованы в качестве сравнительных данных в дальнейших исследованиях), определена их вариативность. Наиболее изменчивыми оказались параметры длительности, безотносительно элемента, к которому они относились.
С учетом размаха изменчивости отдельных элементов, песня выпи неизменна в разных – в том числе удаленных друг от друга – частях ареала. Одинаковыми, не зависящими от географического местоположения, оказываются как общий рисунок начала песни (три «pump»- и один «boom»-слоги), так и физические параметры слогов и их элементов.
Результаты акустических наблюдений могут использоваться при разработке программ мониторинга как непосредственно большой выпи, так и плавневых биотопов в целом.
Ключевые слова: выпь, вокализация, вариабельность
Ianovschi I.Yu.
Postgraduate student, St. Petersburg State University
ON THE DEMONCTRATIVELY VOCALIZATIONS OF GREAT BITTERN (BOTAURUS STELLARIS)
Abstract
This research is devoted to time-and-frequency analysis of variability of great bittern’s calls (Botaurus stellaris). We also provided the profound literature review in this field.
Vocalization of the great bittern is considered to be similar in functions to birdsong of Passerines.
This research was made on the territory of wetlands of Pskov, Russia, during the period of male’s vocal activity. We represented results of parametric measurements and variability of great bittern species specific calls. These results can be used for comparative analysis in a following research. The highest rates of variability were estimated for a duration parameter irrelative to the belonging types.
However, in spite of the high rates of variability of the elements, call structure remains stable within different parts of the highly ranged natural habitat of the species. We defined geographically independent calls similarity as analogous structure of the call’s first parts (three ‘pump’ and one ‘boom’ elements); and physical parameters of the call’s structure and the elements.
The obtained results can be used for future development of species monitoring programs for great bittern as a particular case, and for wetlands in general.
Key words: great bittern, vocalization, variability
Большая или евразийская выпь (Botaurus stellaris) известна своей скрытностью. Ее поведенческие особенности, покровительственная окраска, предпочтения в выборе мест гнездования являются серьезным препятствием при проведении учетов или наблюдений за этой птицей. В то же время, необычайно громкие демонстративные вокализации самца выпи можно уверенно идентифицировать даже на удалении в несколько километров, что позволяет заменить традиционные визуальные наблюдения наблюдениями акустическими. Исследования причин вариабельности демонстративных вокализаций позволяют лучше понимать состояние отдельных особей и, как следствие, осуществлять более емкий в информационном смысле мониторинг популяции этой скрытной, а во многих странах – редкой птицы.
Звуковые демонстрации самцов выпи направленные на привлечение полового партнера и на маркирование своей территории, в функциональном отношении соответствует песням воробьиных птиц [13, 18]. Следовательно, несмотря на отсутствие благозвучия для человеческого слуха, допустимо эти демонстративные вокализации называть «песней».
История вопроса
Попытки описать песню самца выпи и проанализировать механику звукоизлучения предпринимались давно. Так, в XVIII веке крики выпи описывали как «прерванный рев быка». Громкость криков при этом объясняли явлениями резонанса, рассматривая в качестве резонатора само тело птицы [19]. На рубеже XIX и XX веков появился своеобразный взгляд, согласно которому при вокализации самец выпи погружает клюв в воду и тем достигает необходимого характера звучания своей песни. Тогда же появилось иное – более рациональное – объяснение, указывающее на способность пищевода выпи к сильному растяжению и к наличию на стенках пищевода особой мускулатуры, способствующей громкой вокализации [5]. Анатомические исследования американской выпи (Botaurus lentiginosus), имеющей сходную вокализацию, и наблюдения на живых особях евразийской выпи, в целом, укрепили предположение о ключевой роли верхних отделов пищевода в формировании громких демонстративных криков [8, 18]. Шейный отдел самца выпи претерпевает изменения в течение года и весной наблюдается увеличение объема мускулатуры шеи, что рассматривается, как адаптация к громкой демонстративной вокализации [1, 18].
Работы, в том или ином виде посвященные собственно вокализации выпи, можно разделить на три группы: 1) анализ песни как таковой, 2) анализ изменчивости песенной активности самцов выпи, 3) изучение экологии вида с применением акустического метода учета вокализирующих самцов.
Работ, относимых к третьей группе, вероятно, более всего, но прямого отношения к изучению демонстративных вокализаций выпи они не имеют.
До введения методик спектрального анализа, описания песни выпи носили словесный, звукоподражательный характер. Вероятно, одно из первых наиболее полных описаний песни выпи с применением физических терминов и понятий, предложено P.K. McGregor и коллегами в двух работах [12, 9]. Авторы указали на наличие в песне двух типов слогов, отличных по своим физическим характеристикам – негромких, похожих на вдох «pump»-слогов и громких, хорошо слышимых на больших расстояниях «boom»-слогов. Кроме того, было показано, что «boom»-слоги состоят из двух элементов – первого и главного – при чем, первый элемент аналогичен «pump»-слогу, а главный элемент (наиболее громкий в песне выпи) может издаваться птицей в искаженном виде (т.н. «poor boom»-слоги). Введенная ими терминология и структурное деление песни в дальнейшем сохранялись всеми авторами, работающими по данной теме. В обеих работах исследовался вопрос о возможности индивидуальной идентификации самцов выпи по их песням. Было показано, что между отдельными особями существуют отличия, отраженные в длительности и основной частоте «boom»-слогов. Однако, основное внимание в работах было уделено вариабельности, связанной с изменением числа «boom»- и «poor boom»-слогов в песне, т.к. эти характеристики легко уловимы «на слух» и не требуют использования специальной аппаратуры. Дополненные в дальнейшем обобщающей статьей по британской популяции [10], эти работы показали принципиальное наличие индивидуальных черт в песне самца выпи. И хотя, при сравнении в пределах одного временного среза, самцы по этому параметру достоверно различались, при анализе вокализаций, взятых за весь период вокальной активности, достоверность идентификации особей резко падала. Таким образом, была выявлена не только высокая индивидуальная, но и межсезонная вариабельность песен выпи по числу слогов с типичной и атипичной физической структурой.
Функциональная организация песни выпи была проанализирована L. Puglisi и коллегами [18]. Согласно их наблюдениям, корреляции между частотно-временными характеристиками отдельных компонентов песни выражены слабо и, следовательно, у выпи отсутствует твердая структура песни. Главную роль в коммуникации между особями, возможно, играют громкие элементы песни (главные элементы «boom»-слогов), так как они в качественном и количественном отношении оказались наиболее изменчивыми. В то же время была отвергнута идея о возможности различения особей, определения их физиологического состояния по числу «boom» и «poor boom»-слогов в песне. Вслед за Voisin [20] авторы работы высказали предположение, что «pump»-слоги и первый элемент «boom»-слогов – т.е. элементы, напоминающие звук вдоха – на самом деле связаны с актом дыхания. Следовательно, эти звуки можно считать техническими, не имеющими прямого отношения к коммуникации.
Дальнейшие исследования, проведенные на примере двух итальянских популяций [16], показали, что параметры длительности оказались более надежным ключом при индивидуальном распознавании поющих самцов, чем параметры частоты. В этой связи основная роль во внутривидовой коммуникации выпи была отведена длительности отдельных элементов или сочетанию частота-длительность.
Изменения в вокализации самцов выпи на протяжении сезона размножения, а также в зависимости от социальных и климатических факторов подробно изучались на примере популяции, обитающей в парке «Camargue», Южная Франция [14, 15] и на рыборазводных прудах в районе Люблина (Польша) [13]. Согласнорезультатам наблюдений, средняя длительность песен менялась таким образом, что в течение марта она (выраженная в количестве слогов) сокращалась, а с начала апреля вновь нарастала, достигая максимума в первой половине мая. При этом в марте высока была доля «poor boom»-слогов. Заметное влияние на интенсивность вокализаций оказывали абиотические факторы – ветер, температура воздуха, осадки. Были показаны достоверные различия в характере вокализаций самцов, находящихся в зоне акустического контакта и акустически изолированных.
Общее изменение акустической активности самцов выпи на протяжении суток показаны на примерах популяций Псковского Поозерья [2, 3, 4] и дельты р. Роны [14]. В общем случае показано, что пик звуковой активности приходится на сумерки после захода солнца и перед его восходом. Утренний пик преобладает над вечерним, при том, что в разгар сезона голосовой активности высока интенсивность пения, в том числе, и в дневные часы.
Суммируя, следует отметить, что наиболее полно исследован вопрос о изменении интенсивности вокализации самцов выпи в зависимости от разных внешних факторов. Так же, достаточно подробно показана высокая индивидуальная вариабельность отдельно взятых элементов песни. Особенно большое внимание исследователей уделено изменениям в соотношении «boom» и «poor boom»-слогов.
В то же время нельзя не отметить два момента: малое, в целом, количество работ, посвященных непосредственно песне, и ограниченность территории, на которой проводились исследования.
Основная масса работ выполнена на изолированных популяциях в двух географических локусах – в Англии и в северном Средиземноморье (Прованс, Западная Италия). Средиземноморская популяция в районах проведения наблюдений не только размножается, но и гнездится, в Англии гнездовой и зимний ареал перекрываются мало, но при этом имеется сильная нарушенность естественных местообитаний и, как следствие, очень низкая плотность вида. Нарушение местообитаний и сильный антропогенный пресс (что в полной мере относится и к средиземноморскому району исследований) оказывают корректирующее воздействие на поведение выпи [17], что несколько снижает возможности экстраполяции на вид в целом полученных данных. В частности, результаты изучения вокализации выпи в Италии плохо укладывались в выводы, полученные на основании изучения английской популяции [16]. С другой стороны, результаты исследований выпи в Польше [13] вполне согласовались с данными по средиземноморской популяции (при этом стоит учитывать, что работы в Польше велись не в естественных биотопах, но на рыборазводных прудах).
Таким образом, для лучшего понимания закономерностей, влияющих на поведение большой выпи в целом и на ее демонстративные вокализации в частности, необходимо расширение ареала исследований. Особенно важным представляется проведение исследований в местах с сохранившимися естественными биотопами и нормальной плотностью вида. Это позволило бы лучше трактовать ситуативную изменчивость вокализаций самцов выпи и, как следствие, успешнее проводить природоохранные мероприятия.
Материал и методы
Акустические наблюдения за голосовой активностью выпи проводились, с перерывами, в 2008 - 2013 гг. на территории Национального парка «Себежский» и в его окрестностях. Всего, наблюдения велись за 19 самцами.
Работы проводились на озерах Ороно, Себежское, Осыно, на болоте Красиковское, в пойме реки Нища. Дополнительно были привлечены данные наблюдений с северного Приладожья (окрестности пос. Импилахти, Республика Карелия) и материалы из фонотеки кафедры зоологии позвоночных Санкт-Петербургского государственного университета. Все фонотечные записи были сделаны в период с 1980 по 2000 года в Псковском Поозерье.
Запись поющих самцов выпи производилась на магнитофон «Marantz» PMD-222, обладающий динамическим диапазоном 40-12500 Гц, с использованием направленного конденсаторного микрофона «Audio-technica» АТ815В 460 мм с динамическим диапазоном 30-20000 Гц, в апреле-июне, преимущественно в период вечерних сумерек.
Выделение структурных единиц песни основывалось на описании, приведенном в ранних исследованиях по данной тематике [9, 12]. Измерение физических параметров осуществлялось в программе «Praat», версии 5143 для Windows, разработки Paul Boersma и David Weenink.
Математическая обработка собранных данных осуществлялась в программе «Statistica 7.1».
Структура песни
Песня большой выпи слагается из последовательности т.н. «boom»-слогов, каждый из которых состоит из двух элементов – первого и главного. Первый элемент на слух воспринимается, как звучный «вдох», главный элемент – как мощный, гулкий «выдох». Первый «boom»-слог состоит только из одного – главного – элемента, при этом имеет в своем начале короткий низкочастотный участок. «Boom»-слоги могут издаваться в несколько измененном виде – более короткие и менее структурированные – для которых применяется обозначение «poor boom»-слогов.
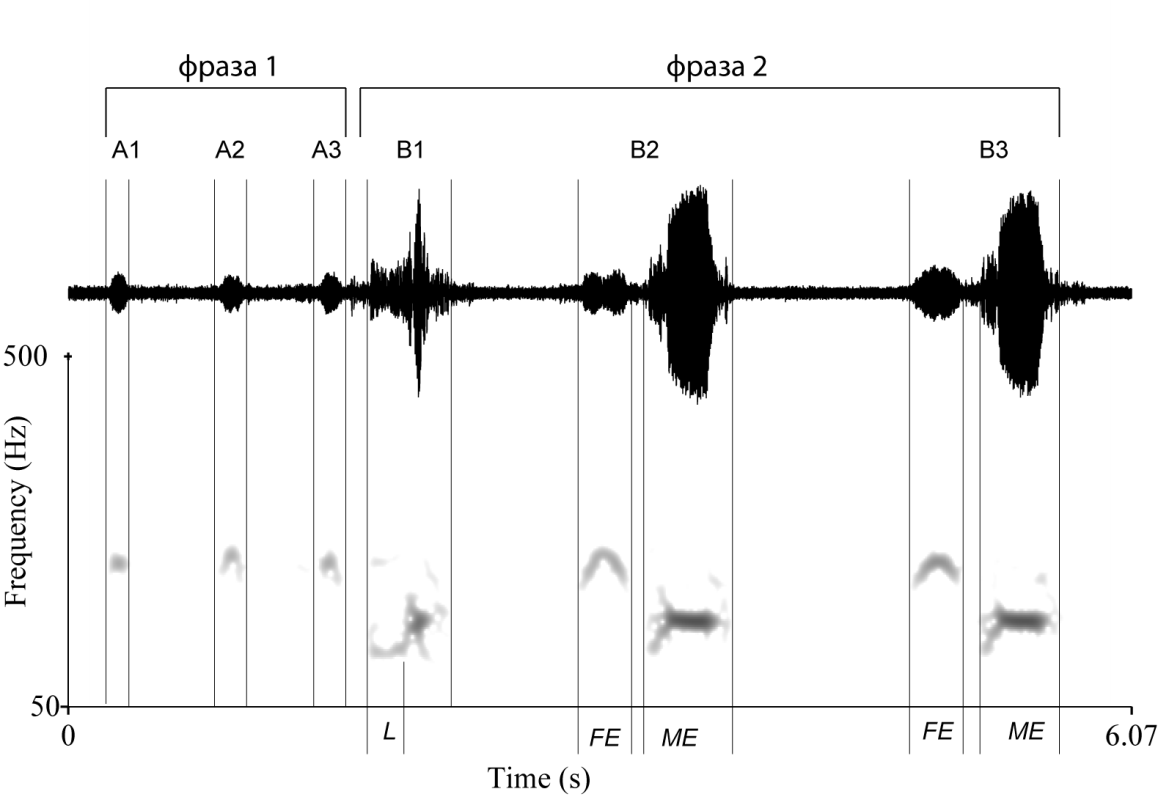
Рис.1 Амплитудная (вверху) и спектральная (внизу) формы образца песни самца выпи. А1 – А3 – т.н. «pump»-слоги, В1 – В3 – т.н. «boom»-слоги (В1 имеет измененную структуру и относится к подтипу «poor boom»-слогов). L – низкочастотный участок в начале главного элемента первого в песне «boom»-слога; FE – первый элемент «boom»-слога (отсутствует у В1); ME – главный элемент «boom»-слога.
Серия «boom»-слогов предваряется короткой последовательностью относительно тихих, также напоминающих вдох, слогов, которые принято обозначать, как «pump»-слоги [9, 12]. Кроме того, могут присутствовать щелчки клювом [1, 21], которые или предшествуют «pump»-слогам, или сопутствуют им. В какой мере они обязательны при демонстративной вокализации выпи – неизвестно.
Принимая во внимание классификацию, предложенную C.K.Catchpole и P.J.R. Slater [7], песню самца выпи можно разделить на две фразы (рис.1).
Результаты
Согласно нашим наблюдениям, песня выпи может содержать от 1 до 11 «boom»-слогов, но чаще их от 4 до 6. Длительность слога, с учетом обоих элементов, составляет почти секунду. Разница между звуковой мощностью обоих элементов - 22,2 дБ (n=29, sx=1,98). Длительность паузы между элементами внутри слога - 0,13с (n= 166, sx=0,1).
Огибающая интенсивности главного элемента «boom»-слогов имеет простую Λ-образную форму, в начале элемента усложненную дополнительной амплитудной модуляций, за счет которой формируется дополнительный амплитудный пик. Разница между дополнительным и основным амплитудными пиками составляет 7,7 дБ (n=30, sx=2,8). Разница между амплитудным максимумом первого «boom»-слога и предваряющего его низкочастотного участка – 14,8 дБ (n=15, sx=1,6).
Первая фраза, в тех случаях, когда качество аудиоматериала позволяло уверенно идентифицировать слоги, всегда состояла из трех слогов.
Огибающая интенсивности первых двух «pump»-слогов имеет Λ-образную форму, огибающая третьего слога и первых элементов «boom»-слогов – Μ-образную форму. Разница между максимумом и минимумом звуковой энергии в пределах этого элемента - 6,1 дБ (n=29, sx=1,5).
Частотный диапазон песни в целом лежит в полосе от 95 до 600-700 Гц, если рассматривать слоги с учетом гармонических составляющих. Если же принимать во внимание только наиболее энергоемкую полосу, тогда верхняя граница составляет 220 Гц. При этом наиболее высокочастотными оказываются «pump»-слоги и первые элементы «boom»-слогов.
Частотная модуляция «pump»-слогов и первого элемента «boom»-слогов имеет Λ-образную форму и спектральный максимум приходится на участок с наименьшей амплитудой.
Частота в начале главного элемента «boom»-слога, в общем случае, повышается, после перегиба в точке спектрального максимума несколько понижается и переходит в «плато». Длительность частотно модулированного участка в пределах главного элемента «boom»-слога составляет 0,2 с (n=147, sx=0,1).
Данные параметрических измерений приведены в таблице 1.
Таблица 1. Средние значения и коэффициент вариации отдельных элементов демонстративных вокализаций выпи
|
Элемент |
N |
среднее |
ст. откл. |
V, % |
|
Длительность 1-го «pump»-слога, с |
45 |
0,11 |
0,03 |
32 |
|
Длительность 2-го «pump»-слога, с |
51 |
0,15 |
0,05 |
33 |
|
Длительность 3-го «pump»-слога, с |
51 |
0,19 |
0,1 |
48 |
|
Длительность низкочастотного участка в начале 1-го «boom»-слога, с |
92 |
0,19 |
0,03 |
15 |
|
Длительность первого элемента 2-го «boom»-слога, с |
83 |
0,33 |
0,1 |
33 |
|
Длительность первого элемента 3-го «boom»-слога, с |
75 |
0,3 |
0,08 |
29 |
|
Длительность главного элемента 1-го «boom»-слога (без учета предваряющего его низкочастотного участка), с |
50 |
0,43 |
0,09 |
18 |
|
Длительность главного элемента 2-го «boom»-слога, с |
84 |
0,54 |
0,11 |
15 |
|
Длительность главного элемента 3-го «boom»-слога, с |
87 |
0,56 |
0,11 |
17 |
|
Спектральный максимум 1-го «pump»-слога, Гц |
44 |
198 |
37,4 |
15 |
|
Спектральный максимум 2-го «pump»-слога, Гц |
50 |
206 |
35 |
10 |
|
Спектральный максимум 3-го «pump»-слога, Гц |
52 |
209 |
28,5 |
5 |
|
Спектральный максимум первого элемента 2-го «boom»-слога, Гц |
81 |
205 |
35,1 |
12 |
|
Спектральный максимум первого элемента 3-го «boom»-слога, Гц |
76 |
206 |
30,3 |
10 |
|
Частота низкочастотного участка в начале 1-го «boom»-слога, Гц |
93 |
119 |
6,6 |
6 |
|
Спектральный максимум главного элемента 1-го «boom»-слога, Гц |
50 |
149 |
10,9 |
6 |
|
Спектральный максимум главного элемента 2-го «boom»-слога, Гц |
83 |
152 |
11,8 |
6 |
|
Спектральный максимум главного элемента 3-го «boom»-слога, Гц |
86 |
151 |
12,5 |
7 |
|
Частота на участке «плато» главного элемента 1-го «boom»-слога, Гц |
49 |
141 |
12,6 |
7 |
|
Частота на участке «плато» главного элемента 2-го «boom»-слога, Гц |
84 |
142 |
13,8 |
6 |
|
Частота на участке «плато» главного элемента 3-го «boom»-слога, Гц |
86 |
141 |
13,3 |
6 |
Обсуждение
Результаты параметрических измерений демонстративных вокализаций самцов выпи, полученные на популяции птиц, обитающих в окрестностях Национального парка «Себежский», почти полностью совпали с данными, приведенными в работе L.Puglisi и коллег [18] для популяции Западной Италии (таб.2).
Таблица 2. Параметрические значения элементов песни выпи двух популяций
|
элемент |
Puglisi et al, 2001 |
наши данные |
||
|
знач. |
ст.откл. |
знач. |
ст.откл. |
|
|
длительность гл. элемента «boom»-слога, с |
0,54 |
0,110 |
0,561 |
0,113 |
|
спектральный максимум гл. элемента «boom»-слога, Гц |
175 |
21,3 |
150,5 |
11,9 |
|
длительность «pump»-слогов, с |
0,14 |
0,041 |
0,154 |
0,074 |
|
спектральный максимум «pump»-слогов, Гц |
206 |
10,6 |
204,8 |
33,7 |
|
длительность первого элемента «boom»-слога, с |
0,39 |
0,086 |
0,317 |
0,093 |
|
спектральный максимум первого элемента «boom»-слога, Гц |
203 |
11 |
205,1 |
32,8 |
Такое совпадение говорит о том, что демонстративные вокализации самцов очень устойчивы на уровне вида. Те отличия, которые все же имеются, по-видимому, объясняются разными методическими приемами при проведении измерений. Вероятно, по этой же причине указывается разное количество слогов в первой фразе. По данным литературы, их бывает от одного до шести [12], но по нашим наблюдениям, их всегда три. Вероятно, в данном случае присутствует системная ошибка: на спектрограмме из-за высокого уровня фоновых шумов один «pump»-слог интерпретируется, как несколько более коротких. Так, например, третий «pump»-слог, имея М-образную амплитудную модуляцию, при увеличении дистанции звукозаписи воспринимается, как два отдельных коротких слога. Если это так, тогда можно говорить о неизменном рисунке начала песни выпи: три слога первой фразы и минимум один слог второй фразы. Это – минимальная звуковая композиция, которая уже может быть воспринята, как демонстративная вокализация самца. Увеличение количества «boom»-слогов зависит от ситуативной и индивидуальной изменчивости. Согласно наблюдениям в окрестностях Парка «Себежский», количество «boom»-слогов может достигать 11 в одной песне, что несколько больше, чем указывается для европейской популяции [12, 18, 16].
В условиях естественных местообитаний в северной части ареала самцы выпи занимают участки плавней на значительном удалении друг от друга. Расстояние от одного самца до другого, как правило, составляет многие сотни метров и может превышать километр. При этом определяющим фактором, по всей видимости, является именно определенная удаленность соседствующих самцов, а не наличие подходящих мест гнездовий. Так, весной, в период формирования индивидуальных участков (который начинается сразу, как только с водоема сходит лед), самцы выпи ревностно отстаивают очень большие площади прибрежных плавней [6]. На озере Осыно, согласно многолетним наблюдениям, выпь может гнездиться в разных местах, но никогда не занимает их все сразу (устное сообщение И.В. Ильинского). В среднем, в районе Псковского Поозерья один вокализирующий самец приходится на 6 – 6,5 км2 акватории [2].
При распространении на такие расстояния и с учетом звукопоглощающего эффекта густой растительности тростниковых плавней [11], тихие элементы песни в значительной степени затухают или могут вовсе теряться в естественном уровне природного шума. В таком случае дистантная коммуникация между соседствующими особями обеспечивается главными элементами «boom»-слогов. Первый «boom»-слог предваряется низкочастотным участком, который значительно тише основной громкости слога и тоже может маскироваться средой. Без его учета, первый слог второй фразы оказывается достоверно короче, чем все последующие слоги (рис.2).
На первый взгляд, эти данные противоречат данным по итальянской популяции, в ходе которых не удалось выявить каких-либо закономерных отличий в длительности «boom»-слогов [18]. Однако, причина различий может объясняться разными методическими приемами при проведении исследований. Длительность первого слога второй фразы, принятая с учетом предваряющего его низкочастотного участка, достоверно больше длительности главных элементов последующих «boom»-слогов (рис.2, левый график). Но низкочастотный участок хорошо маскируется природным шумом при увеличении расстояния до вокализирующей птицы. При его включении в измерение длительности слога и при разных условиях звукозаписи, итоговые значения будут отражать как индивидуальную изменчивость вокализаций, так и акустические условия проведения аудиозаписи.
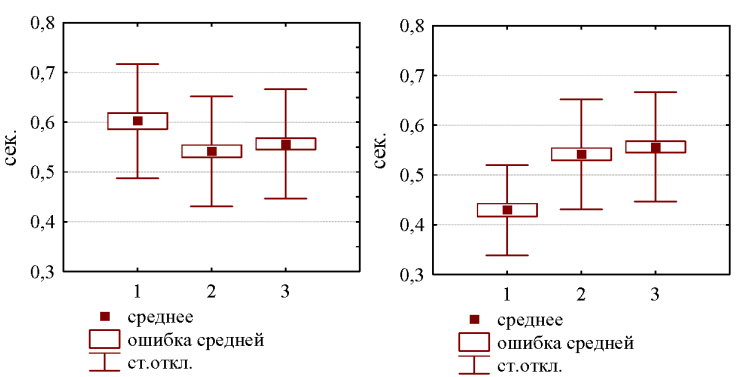
Рис.2 Длительность главных элементов первых трех (1-3) «boom»-слогов (на левом графике первый слог принят с учетом длительности низкочастотного участка, на правом графике представлена длительность только главных элементов, без учета низкочастотного участка, предваряющего первый «boom»-слог).
Заключение
Опираясь на анатомические данные представителей рода Botaurus [8, 18], в настоящее время сложилось устойчивое представление о том, что громкость главных элементов «boom»-слогов в демонстративных вокализациях выпи обеспечивается за счет резонансных явлений. Остальным элементам песни (первые элементы «boom»-слогов, «pump»-слоги) в этом случае отводится роль своего рода технических звуков, связанных с процессом дыхания и не играющим важной роли в процессе внутривидовой коммуникации [18, 20].
Не обсуждая коммуникативную нагрузку разных элементов песни, стоит отметить, что параметры частоты (безотносительно элемента, с которым они были связаны) оказались менее вариабельны, чем параметры длительности. Наиболее изменчивыми оказались «pump»-слоги и первые элементы «boom»-слогов.
Демонстративная вокализация, как поведенческая реакция самца выпи в период гнездования, оказалась очень устойчивой и не зависящей от географического местоположения. Неизменность проявилась как на уровне общего рисунка (обязательное начало: три «pump»-слога и один «boom»-слог), так и на уровне физических параметров отдельных элементов. Различия найдены в максимальной продолжительности песни, но речь, по-видимому, должна идти только об индивидуальной изменчивости, а не о каких-либо географических вариациях. К таким же постоянным, не зависящим от географии, особенностям следует отнести и найденные отличия в длительности первого и последующих «boom»-слогов (точнее – их наиболее громких элементов).
Автор выражает благодарность И.В. Ильинскому и С.А. Фетисову за всестороннюю помощь в проведении исследований.
Список литературы
Дмитренок М. Г. Выпь Botaurus stellaris (Linnaeus, 1758) // Птицы России и сопредельных регионов. Пеликанообразные, аистообразные, фламингообразные. - М.: Товарищество научных изданий КМК. – 2011. - С. 155-177.
Ильинский И. В., Пчелинцев В. Г., Фетисов С. А. Наблюдения за гнездованием выпи Botaurus stellaris в Себежском Поозерье (Псковская область) // Русск. орнитол. журн. – 1997. Экспресс-выпуск. № 17. - С. 16-21.
Ильинский И. В., Фетисов С. А. Характеристика демонстративного "пения" большой выпи // Изучение птиц СССР, их охрана и рациональное использование: тезисы докл. I съезда Всесоюзн. орнитол. общ-ва и IX Всесоюзн. орнитол. конф. - Л.: Изд-во АН СССР, 1986. - С. 264-266.
Ильинский И. В., Фетисов С. А., Яновский И. Ю. Сезонные и суточные ритмы пения выпи Botaurus stellaris в условиях Псковского Поозерья // Русск. орнитол. журн. – 2010. Экспресс-выпуск. № 618. - С. 2207-2211.
Холодковский Н. А., Силантьев А. А. Птицы Европы. Практическая орнитология с атласом европейских птиц. - СПб, Изде А.Ф. Девриена, 1901. – 635 с.
Яновский И. Ю. К вопросу о формировании индивидуальных территорий самцами выпи Botaurus stellaris // Русск. орнитол. журн. – 2012. Экспресс-выпуск. № 720. - С. 117-124.
Catchpole C. K., Slater P. J. R.. Bird song: biological themes and variations. Second edition. - Cambridge Univ.Press., 2008 - 335 p.
Chapin J. P. The function of the oesophagus in the bittern’s booming // The Auk. – 1922. Vol. 39. - P.196-202.
Gilbert G., McGregor P. K., Tyler G. A. Vocal individuality as a census tool: Practical considerations illustrated by a study of two rare species // Journal of Field Ornithology. – 1994. Vol. 65. - P. 335-348.
Gilbert G., Tyler G. A., Smith K. W. Local annual survival of booming male Great Bittern Botaurus stellaris in Britain, in period 1990-1999 // Ibis. – 2002. Vol. 144. - P. 51-61.
Ianovschi I., Osipova L. Sound insulation of artificial and natural sound signals in reeds habitats // ACOUSTICS’08: Abstracts. – Paris, 2008. - P. 1454
McGregor P. K., Byle P. Individual distinctive bittern booms: potential as a census tool // Bioacoustic. – 1992. Vol. 4. – P. 93-109.
Polak M. Booming activity of male Great Bittern Botaurus stellaris in relation to reproductive cycle and harem size // Ornis Fennica. – 2006. Vol. 83. - P. 27-33.
Poulin B., Lefebvre G. Optimal sampling of booming Bitterns Botaurus stellaris // Ornis Fennica. – 2003a. Vol. 80. – P. 11-20.
Poulin B., Lefebvre G. Variation in booming among Great Bitterns Botaurus stellaris in the Camargue, France // Ardea. – 2003b. Vol. 91, № 2. – P. 177-181.
Puglisi L., Adamo C. Discrimination of individual voices in male Great Bittern (Botaurus stellaris) in Italy // The Auk. – 2004. Vol. 121, № 2. P. 541-547.
Puglisi, L., Adamo C., Baldaccini, N. E., 2005. Man-induced habitat changes and sensitive species: a GIS approach to the Eurasian Bittern (Botaurus stellaris) distribution in a Mediterranean wetland // Biodiversity and Conservation. – 2005. Vol. 14. - P. 1909-1922.
Puglisi, L., Pagni, M., Bulgarelli, Ch., Baldaccini, N. E. The possible functions of calls organization in the bittern (Botaurus stellaris) // Italian Journal of Zoology. – 2001. Vol. 68. – P. 315–321.
Studer J.H. The birds of North America. - N.Y., 1903. - 191 p.
Voisin C. The herons of Europe. - Bloomsbury Publishing PLC, 1991. - 376 p.
Zimmermann R. Zur Oekologie und Biologie der Grogen Rohrdommel, Botanrus stellaris L., in der Oberlausitzer Niederung. // Journal of Ornithology. - 1929. Vol. 77, №4. – P. 249-266.
Список литературы
Дмитренок М. Г. Выпь Botaurus stellaris (Linnaeus, 1758) // Птицы России и сопредельных регионов. Пеликанообразные, аистообразные, фламингообразные. - М.: Товарищество научных изданий КМК. – 2011. - С. 155-177.
Ильинский И. В., Пчелинцев В. Г., Фетисов С. А. Наблюдения за гнездованием выпи Botaurus stellaris в Себежском Поозерье (Псковская область) // Русск. орнитол. журн. – 1997. Экспресс-выпуск. № 17. - С. 16-21.
Ильинский И. В., Фетисов С. А. Характеристика демонстративного "пения" большой выпи // Изучение птиц СССР, их охрана и рациональное использование: тезисы докл. I съезда Всесоюзн. орнитол. общ-ва и IX Всесоюзн. орнитол. конф. - Л.: Изд-во АН СССР, 1986. - С. 264-266.
Ильинский И. В., Фетисов С. А., Яновский И. Ю. Сезонные и суточные ритмы пения выпи Botaurus stellaris в условиях Псковского Поозерья // Русск. орнитол. журн. – 2010. Экспресс-выпуск. № 618. - С. 2207-2211.
Холодковский Н. А., Силантьев А. А. Птицы Европы. Практическая орнитология с атласом европейских птиц. - СПб, Изде А.Ф. Девриена, 1901. – 635 с.
Яновский И. Ю. К вопросу о формировании индивидуальных территорий самцами выпи Botaurus stellaris // Русск. орнитол. журн. – 2012. Экспресс-выпуск. № 720. - С. 117-124.
Catchpole C. K., Slater P. J. R.. Bird song: biological themes and variations. Second edition. - Cambridge Univ.Press., 2008 - 335 p.
Chapin J. P. The function of the oesophagus in the bittern’s booming // The Auk. – 1922. Vol. 39. - P.196-202.
Gilbert G., McGregor P. K., Tyler G. A. Vocal individuality as a census tool: Practical considerations illustrated by a study of two rare species // Journal of Field Ornithology. – 1994. Vol. 65. - P. 335-348.
Gilbert G., Tyler G. A., Smith K. W. Local annual survival of booming male Great Bittern Botaurus stellaris in Britain, in period 1990-1999 // Ibis. – 2002. Vol. 144. - P. 51-61.
Ianovschi I., Osipova L. Sound insulation of artificial and natural sound signals in reeds habitats // ACOUSTICS’08: Abstracts. – Paris, 2008. - P. 1454
McGregor P. K., Byle P. Individual distinctive bittern booms: potential as a census tool // Bioacoustic. – 1992. Vol. 4. – P. 93-109.
Polak M. Booming activity of male Great Bittern Botaurus stellaris in relation to reproductive cycle and harem size // Ornis Fennica. – 2006. Vol. 83. - P. 27-33.
Poulin B., Lefebvre G. Optimal sampling of booming Bitterns Botaurus stellaris // Ornis Fennica. – 2003a. Vol. 80. – P. 11-20.
Poulin B., Lefebvre G. Variation in booming among Great Bitterns Botaurus stellaris in the Camargue, France // Ardea. – 2003b. Vol. 91, № 2. – P. 177-181.
Puglisi L., Adamo C. Discrimination of individual voices in male Great Bittern (Botaurus stellaris) in Italy // The Auk. – 2004. Vol. 121, № 2. P. 541-547.
Puglisi, L., Adamo C., Baldaccini, N. E., 2005. Man-induced habitat changes and sensitive species: a GIS approach to the Eurasian Bittern (Botaurus stellaris) distribution in a Mediterranean wetland // Biodiversity and Conservation. – 2005. Vol. 14. - P. 1909-1922.
Puglisi, L., Pagni, M., Bulgarelli, Ch., Baldaccini, N. E. The possible functions of calls organization in the bittern (Botaurus stellaris) // Italian Journal of Zoology. – 2001. Vol. 68. – P. 315–321.
Studer J.H. The birds of North America. - N.Y., 1903. - 191 p.
Voisin C. The herons of Europe. - Bloomsbury Publishing PLC, 1991. - 376 p.
Zimmermann R. Zur Oekologie und Biologie der Grogen Rohrdommel, Botanrus stellaris L., in der Oberlausitzer Niederung. // Journal of Ornithology. - 1929. Vol. 77, №4. – P. 249-266.