ИЗМЕНЕНИЯ ЭЭГ В НАЧАЛЬНЫЙ ПЕРИОД ЗАСЫПАНИЯ И ВО ВРЕМЯ ПЕРВОГО ЭПИЗОДА КРАТКОВРЕМЕННОГО ДНЕВНОГО СНА НА ФОНЕ ВЫПОЛНЕНИЯ ПСИХОМОТОРНОГО ТЕСТА

ИЗМЕНЕНИЯ ЭЭГ В НАЧАЛЬНЫЙ ПЕРИОД ЗАСЫПАНИЯ И ВО ВРЕМЯ ПЕРВОГО ЭПИЗОДА КРАТКОВРЕМЕННОГО ДНЕВНОГО СНА НА ФОНЕ ВЫПОЛНЕНИЯ ПСИХОМОТОРНОГО ТЕСТА
Научная статья
Петренко Н.Е.1 , Черемушкин Е.А.2, *, Дорохов В.Б.3
1 ORCID: 0000-0003-3639-8957;
2 ORCID: 0000-0001-6902-8077;
3 ORCID: 0000-0003-3533-9496;
1– 3 Институт высшей нервной деятельности и нейрофизиологии Российской академии наук, Москва, Россия
* Корреспондирующий автор (khton[at]mail.ru)
Аннотация
У 24 здоровых испытуемых исследовали начальный период засыпания и первый эпизод кратковременного дневного сна на фоне выполнения психомоторной деятельности. Анализировали мощностные характеристики ритмов ЭЭГ в 3-х ситуациях: 1 – при деятельности в период полной бдительности, в самом начале опыта, 2 – при деятельности перед первым эпизодом кратковременного сна, 3 – при кратковременном (от 30 до 60 с) сне. Показано, что непосредственно перед эпизодом сна на фоне роста дельта- и тета-колебаний сохраняется определенный уровень активации переднецентральных областей коры мозга, что позволяет выполнять психомоторную деятельность в этот период. Во время эпизода дневного сна (только 1-ой стадии, не переходящей во 2-ую) эта активация, характеризующаяся альфа- и низкочастотными бета-колебаниями, сохраняется, что позволяет быстро проснуться и вернуться к деятельности, прерванной сном. Обнаруженные нейрокорреляты ранней стадии засыпания и эпизода кратковременного сна на фоне выполнения психомоторного теста позволяют оценить функциональное состояние и уровень сознания субъекта при выполнении длительной монотонной трудовой деятельности.
Ключевые слова: засыпание, психомоторная деятельность, кратковременный дневной сон, ЭЭГ.
CHANGES IN THE EEG DURING THE INITIAL PERIOD OF FALLING ASLEEP AND THE FIRST EPISODE OF SHORT-TERM DAYTIME SLEEP DURING A PSYCHOMOTOR TEST
Research article
Petrenko N.E.1 , Cheremushkin E.A.2, *, Dorokhov V.B.3
1 ORCID: 0000-0003-3639-8957;
2 ORCID: 0000-0001-6902-8077;
3 ORCID: 0000-0003-3533-9496;
1– 3 Institute of Higher Nervous Activity and Neurophysiology of the Russian Academy of Sciences, Moscow, Russia
*Corresponding author (khton[at]mail.ru)
Abstract
In 24 healthy subjects, the initial period of falling asleep and the first episode of short-term daytime sleep were studied during psychomotor activity. The power characteristics of EEG rhythms were analyzed in 3 situations: 1 – during activity while being fully awake, at the very beginning of the experiment, 2 – during activity before the first episode of short–term sleep, 3 – during short (from 30 to 60 seconds) sleep. It is shown that immediately before the episode of sleep, against the background of the growth of delta and theta oscillations, a certain level of the anterocentral regions of the cerebral cortex remains, which allows performing psychomotor activity during this period. During an episode of daytime sleep (only the 1st stage, which does not pass into the 2nd), this activation, characterized by alpha and low-frequency beta fluctuations, persists, which allows to wake up quickly and return to activities interrupted by sleep. The detected neurocorrelates of the early stage of falling asleep and an episode of short-term sleep against the background of a psychomotor test allows to assess the functional state and level of consciousness of the subject during a long monotonous activity.
Keywords: falling asleep, psychomotor activity, short-term daytime sleep, EEG.
Введение
Засыпание – это переходный процесс между двумя состояниями сознания: бодрствованием и сном. При засыпании теряется способность к восприятию внешней среды, к осуществлению поведенческих актов и происходит постепенное снижение уровня сознания [1, С. 60]. В ЭЭГ оно сопровождается переходом от колебаний биопотенциалов высокой частоты и низкой амплитуды к низкочастотным, высокоамплитудным медленным волнам и т. н. “сонным” веретенам, и представляет собой постепенный, неоднородный процесс, как с пространственной, так и с временно́й точки зрения [2, С. 22]. В разных областях коры головного мозга во время наступления сна в одно и то же время наблюдалась биоэлектрическая активность, встречающаяся как при бодрствовании, так и во сне [3, С. 33], [4, С. 12–13]. При этом до сих пор не существует единой точки зрения, что считать началом сна [3, С. 25]. Большинство исследователей определяют его с момента появления в ЭЭГ «сонных» веретен и К-комплексов, т.е. с началом 2-ой стадии сна. Значительно реже за начало сна принимают появление признаков 1-ой стадии сна.
Целью работы было изучение динамики биоэлектрической активности коры головного мозга при засыпании и на 1-ой стадии кратковременного дневного сна. В задачу входило сопоставление мощностных характеристик ЭЭГ
в дельта-, тета-, альфа- и сигма-диапазонах при засыпании на фоне выполнения психомоторной деятельности.
Методы и принципы исследования
Испытуемые. 34 человека (26 женщин и 8 мужчин, возраст 19–22 года), практически здоровые люди, правши, студенты московских вузов. Все были ознакомлены с процедурой опыта и дали согласие на участие в нем. Исследование соответствовало этическим нормам Хельсинской декларации Всемирной медицинской ассоциации “Этические принципы проведения научных медицинских исследований с участием человека” с поправками 2000г.
Процедура исследования. Время эксперимента: с 13.00 до 16.00. Продолжительность – от 55 мин до 1 часа 10 мин. Испытуемый располагался на кушетке в затемненном и звукоизолирующем помещении. После инструкции он выполнял непрерывно-дискретный психомоторный тест. Испытуемый считал «про себя» от 1 до 10, при этом одновременно при каждом счете он нажимал на кнопку большим пальцем правой руки (1 фаза теста), которая была закреплена на указательном пальце. Далее он продолжал считать «про себя» от 1 до 10, но уже без нажатий (2 фаза). Монотонный характер теста вызывает быстрое снижение уровня бодрствования с множественными кратковременными эпизодами засыпания―пробуждения в течение одного часа эксперимента [5, С. 1233–1234].
Регистрировали ЭЭГ от 17 отведений, расположенных в соответствии со схемой 10–20% (F3, F4, F7, F8, Fz, C3, C4, Cz, T3, T4, P3, P4, Pz, T5, T6, O1, O2). Отведение было монополярным, референтный электрод – объединенный ушной. Также записывалась электроокулограмма (ЭОГ), миограмма (ЭМГ) и механограмма нажатий на кнопку. Регистрацию всех показателей проводили с помощью системы Neocortex–Pro («Neurobotics», Россия). Частота дискретизации – 250 Гц. Полоса пропускания частот: 0,5–70 Гц. ЭЭГ регистрировали с помощью шлема с хлорсеребряными электродами с сопротивлением, не превышающим 5 КОм.
Отбор и анализ данных. Для анализа выбирали 30-секундные отрезки записи ЭЭГ в начале первого эпизода кратковременного сна (1-ая стадия сна, около 1 мин), спустя 10 с после прекращения нажатий на кнопку, без признаков 2-ой стадии сна. Всего отобрали 24 эпизода, по одному у 24 испытуемых. Оценка принадлежности к этому периоду сна, проводилась визуально согласно стандартным критериям AASM [6, С. 16–30]. У этих же испытуемых выделяли 30-секундные отрезки записи ЭЭГ при выполнении психомоторного теста в самом начале эксперимента, в состоянии полной бдительности, и непосредственно перед засыпанием. Для оценки мощностных характеристик биоэлектрической активности коры проводили непрерывное вейвлет-преобразование на основе «материнского» комплексного Morlet-вейвлета (Matlab 78.01, параметры для скриптов из [7, С. 4246]).
Карты распределения значений модуля коэффициента вейвлет-преобразования (КВП) строили в полосе 0,5–16 Гц с шагом 0,5 Гц и разрешением по времени 0,01 с. В дельта- (1–3,5 Гц), тета (4–7,5 Гц), альфа- (8–11,5 Гц) и сигма- (12–16 Гц) спектральных диапазонах проводили усреднение КВП по частоте; далее проводили усреднение по времени внутри 30-секундных временны́х интервалов для всех 3-х исследуемых ситуаций. Для начального эпизода кратковременного сна дополнительно усредняли КВП по времени в 3-х последовательных 10-секундных интервалах. Полученные суммарные амплитудно-мощностные характеристики ЭЭГ логарифмировали и далее анализировали влияние на них различных факторов с помощью дисперсионного анализа (ANOVARM). Проводили два вида анализа. 1. Рассматривали влияние факторов «ситуация»: 3 уровня (деятельность при полной бдительности в самом начале опыта, деятельность перед первым эпизодом кратковременного сна, начало кратковременного сна) и “отведения”: 17 уровней (по числу отведений). 2. Рассматривали влияние факторов «время»: 3 уровня (1–10 с, 11–20 с и 21–30 с исследуемого 30-секундного отрезка ЭЭГ в начале эпизода сна) и «отведения»: 17 уровней (по числу отведений ЭЭГ). Оба анализа осуществляли для каждого из исследуемых частотных диапазонов в отдельности. Все результаты получали с использованием поправки Гринхауса-Гейссера. При помощи парного критерия Стьюдента сопоставляли вычисленные спектральные характеристики ЭЭГ как для 3-х ситуаций 1-го вида анализа, так и для 3-х последовательных 10-секундных интервалов времени для 2-го, в начале эпизода сна. Все статистические вычисления проводили с помощью пакета программ SPSS 13.0.
Основные результаты
Результаты 1-го дисперсионного анализа приведены в Таблице 1.
Таблица 1 – Влияние факторов «ситуация» и «отведение» на мощностные характеристики ЭЭГ
|
Факторы ANOVA RM |
Ритмы |
|||
|
Дельта |
Тета |
Альфа |
Сигма |
|
|
Ситуация |
F(2;40)=6,31 p=0,005 |
F(2;38)=8,92 p=0,001 |
F(1;28)=16,98 p=0,0001 |
F(2;35)=18,71 p=0,0001 |
|
Ситуация* Отведение |
F(6;140)=4,55 p=0,0001 |
F(4;100)=14,29 p=0,0001 |
F(3;70)=10,15 p=0,0001 |
F(4;92)=20,79 p=0,0001 |
Статистическая оценка попарных последовательных изменений в характеристиках ЭЭГ между экспериментальными ситуациями, исследованными в 1-ом виде дисперсионного анализа по отведениям, приведена на рис. 1.
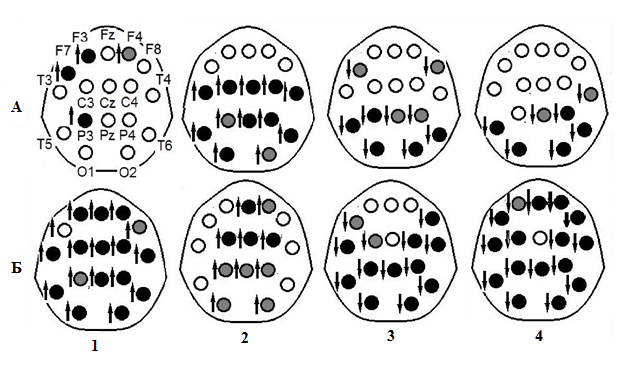
Рис. 1 – Карты-схемы попарных последовательных изменений
в мощностных характеристиках ЭЭГ между экспериментальными ситуациями:
А – от деятельности при полной бдительности, в самом начале опыта, к деятельности перед первым эпизодом кратковременного сна; Б – от деятельности перед 1-ым засыпанием к 1-ому эпизоду кратковременного сна;1–4 – дельта-, тета-, альфа-- и сигма-ритмы соответственно; значки «стрелка вверх» и «стрелка вниз» – увеличение и уменьшение мощности ритмов соответственно; серые и черные круги – значимость изменений: Р < 0,05 и Р < 0,01 соответственно
На карте-схеме в верхнем левом углу рисунка показаны названия отведений ЭЭГ.
Результаты 2-го дисперсионного анализа приведены в Таблице 2.
Таблица 2 – Влияние факторов “время” и “отведение” на мощностные характеристики ЭЭГ
|
факторы ANOVA RM |
ритмы |
|||
|
Дельта |
Тета |
Альфа |
Сигма |
|
|
Время |
|
F(2;22)=5,65 p=0,010 |
F(2;22)=4,37 p=0,025 |
F(2;22)=5,17 p=0,014 |
|
Отведение |
F(3;75)=46,5 p=0,0001 |
F(4;86)=59,9 p=0,0001 |
F(3;90)=91,72 p=0,0001 |
F(4;90)=93,5 p=0,0001 |
|
Время* Отведение |
|
|
F(10;219)=2,34 p=0,013 |
F(9;200)=1,88 p=0,059 |
Анализ динамики мощностных характеристик ЭЭГ в начале сна по отдельным отведениям показал, что существует статистически значимый их рост между 1-ым и 2-ым и 1-ым и 3-им последовательными 10-секундными интервалами времени (рис.2). Различий между 2-ым и 3-им исследуемыми интервалами обнаружено не было.
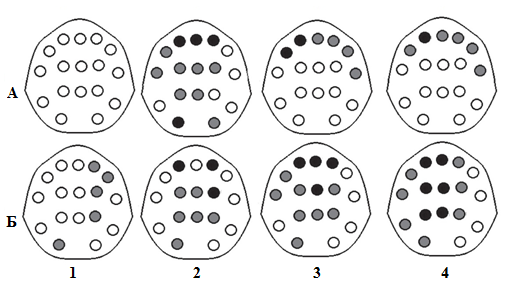
Рис. 2– Карты-схемы статистически значимых изменений (роста) мощностных
характеристик ритмовЭЭГ по отведениям в начале эпизода дневного сна:
А – от 1-го ко 2-ому 10-секундныму отрезку времени сна; Б – от 1-ого к 3-ему; 1–4 – дельта-, тета-, альфа- и сигма-ритмы ЭЭГ соответстивенно; серые и черные круги – P<0,05 и P<0,01 соответственно
Обсуждение
По мере снижения уровня бодрствования от начала опыта к эпизоду кратковременного дневного сна (1-ой стадии), прерывающего выполнение психомоторной деятельности, в большинстве отведений ЭЭГ наблюдается увеличение мощности дельта- и тета-колебаний и снижение альфа- и сигма-колебаний. Эти изменения являются характерными для развития процессов засыпания и последующего погружения в сон с переходом во 2-ую его стадию [3, С. 25], [4, С. 1], [8, С. 99]. При этом отметим: во время деятельности непосредственно перед засыпанием сохраняется уровень активации передних и центральных областей в альфа- и сигма-диапазонах ЭЭГ (рис. 1, А, 3 и рис. 1, А, 4), и даже в начале эпизода сна – в альфа-диапазоне, в дорзо-латеральной префронтальной коре (рис. 1, Б, 3). Мы предполагаем, что при общей тенденции снижения степени бодрствования во время засыпания, сохранение уровня активации лобных областей обеспечивает выполнение в этих условиях психомоторной, когнитивной деятельности. Ее прерывание в начале сна, вероятно, оставляет следовой эффект в виде продолжающейся активации, которая после пробуждения дает возможность быстро возобновить выполнение заданий теста.
Более детальное исследование изменений ЭЭГ в эпизоде кратковременного сна показало рост низкочастотных составляющих ЭЭГ от 1-го к двум последующим 10-секундным интервалам, что указывает на продолжение снижения активации. При этом мы обнаружили увеличение мощностных характеристик в альфа- и сигма-диапазонах, особенно продолжительных в лобных областях отведения (рис. 2, А, 3 и 4; рис 2, Б, 3 и 4). Если учитывать, что сигма-диапазон в нашем исследовании включает в себя высокочастотную альфа- и низкочастотную бета-активность, а при визуальной оценке ЭЭГ сигма-веретена, по частоте совпадающие с этой активностью, отсутствуют, то данный факт, вероятно, отражает рост активации мозговых процессов, которые спустя непродолжительное время приводят к пробуждению. Рост мощности альфа- и бета-ритмов обнаружен при пробуждении из 2-ой стадии как ночного [9, С. 235], [10, С.5196–5197], так и дневного сна [11, С. 215]. В последнем случае данное электрофизиологическое явление опережает начало восстановления психомоторной деятельности и может отражать степень ее эффективности и сопутствующий ей уровень активации сознания.
Заключение
При выполнении психомоторной деятельности непосредственно перед засыпанием в ЭЭГ присутствуют как паттерны биоэлектрической активности, характерные для реализации этой деятельности, так и признаки начальной стадии сна. Сохранение активации в лобных областях мозга в начале эпизода кратковременного дневного сна на 1-ой его стадии позволяет быстро вернуться к выполнению прерванной сном деятельности.
|
Конфликт интересов Не указан. |
Conflict of Interest None declared. |
Список литературы
Tavakoli P. Highly relevant stimuli may passively elicit processes associated with consciousness during the sleep onset period / P. Tavakoli, S. Varma, K. Campbell // Consciousness and Cognition. – 2018. – Vol. 58. – P. 60–74. DOI: 10.1016/j.concog.2017.10.012.
Gorgoni M. Timing and Topography of Sleep Onset: Asynchronies and Regional Changes of Brain Activity / M. Gorgoni, A. D’Atri, S. Scarpelli et al. // Handbook of Behavioral Neuroscience. – 2019. – P. 19–31. DOI:10.1016/b978-0-12-813743-7.00.
Guerrero A.F. Brain dynamics during the sleep onset transition: an EEG source localization study/ A.F. Guerrero, P. Achermann // Neurobiology of Sleep and Circadian Rhythms. – 2018. – Vol. 6. – P. 24–34. DOI: 10.1016/j.nbscr.2018.11.001.
Gorgoni M. The Spatiotemporal Pattern of the Human Electroencephalogram at Sleep Onset After a Period of Prolonged Wakefulness / M. Gorgoni, C. Bartolacci, A. D’Atri et al. // Frontiers in Neuroscience. – 2019. – Vol.13. – P. 1–15. DOI: 10.3389/fnins.2019.00312.
Cheremushkin E.A. Eeg characteristics during short-term spontaneous waking periods of different durations with changes in psychomotor activity induced by falling asleep / E.A. Cheremushkin, N.E. Petrenko, V.B. Dorokhov et al. // Neuroscience and Behavioral Physiology. – 2020. – Vol. 50. – № 9. – P. 1232–1238. DOI: 10.1007/s11055-020-01024-8
Berry R.B. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications, Version 2.2. / R.B. Berry, R. Brooks, C.E. Gamaldo et al. // Darien, Illinois: American Academy of Sleep Medicine, 2015. – 79 p.
Tallon-Baudry C. Induced gamma band activity during the delay of a visual short term memory task in humans / C. Tallon–Baudry, O. Bertrand, F. Peronnet et al. // Journal of neuroscience. – 1998. – Vol .18. – № 11. – P. 4244–4255. DOI: 10.1523/JNEUROSCI.18-11-04244.1998.
Yakovenko I.A. The study of rhythmic component coupling at the first stage of day sleep / I.A. Yakovenko, D.Y. Shumov, N.E. Petrenko et al. // Moscow University Biological Sciences Bulletin. – 2019. – Vol. 74. – № 2. – P. 98-102. DOI: 10.3103/S0096392519020111.
Peter-Derex L. Heterogeneity of arousals in human sleep: A stereo-electroencephalographic study / L. Peter-Derex, M. Magnin, H. Bastuji // Neuroimage. – 2015. – Vol. 123. – P.229–244. DOI: 10.1016/j.neuroimage.2015.07.057.
Ruby P. Dynamics of hippocampus and orbitofrontal cortex activity during arousing reactions from sleep: An intracranial electroencephalographic study / P. Ruby, M. Eskinazi, R. Bouet et al. // Human Brain Mapping. – 2021. – Vol .42. – № 16. – P. 5188-5203. DOI: 10.1002/hbm.25609. doi:10.1002/hbm.25609.
Cheremushkin E.A. Sleep and neurophysiological correlates of activation of consciousness on awakening / E.A. Cheremushkin, N.E. Petrenko, V.B. Dorokhov // Neuroscience and Behavioral Physiology. – 2022. – Vol. 52. – № 2. – P. 213–217. DOI: 10.1007/s11055-022-01226-2.
Список литературы
Tavakoli P. Highly relevant stimuli may passively elicit processes associated with consciousness during the sleep onset period / P. Tavakoli, S. Varma, K. Campbell // Consciousness and Cognition. – 2018. – Vol. 58. – P. 60–74. DOI: 10.1016/j.concog.2017.10.012.
Gorgoni M. Timing and Topography of Sleep Onset: Asynchronies and Regional Changes of Brain Activity / M. Gorgoni, A. D’Atri, S. Scarpelli et al. // Handbook of Behavioral Neuroscience. – 2019. – P. 19–31. DOI:10.1016/b978-0-12-813743-7.00.
Guerrero A.F. Brain dynamics during the sleep onset transition: an EEG source localization study/ A.F. Guerrero, P. Achermann // Neurobiology of Sleep and Circadian Rhythms. – 2018. – Vol. 6. – P. 24–34. DOI: 10.1016/j.nbscr.2018.11.001.
Gorgoni M. The Spatiotemporal Pattern of the Human Electroencephalogram at Sleep Onset After a Period of Prolonged Wakefulness / M. Gorgoni, C. Bartolacci, A. D’Atri et al. // Frontiers in Neuroscience. – 2019. – Vol.13. – P. 1–15. DOI: 10.3389/fnins.2019.00312.
Cheremushkin E.A. Eeg characteristics during short-term spontaneous waking periods of different durations with changes in psychomotor activity induced by falling asleep / E.A. Cheremushkin, N.E. Petrenko, V.B. Dorokhov et al. // Neuroscience and Behavioral Physiology. – 2020. – Vol. 50. – № 9. – P. 1232–1238. DOI: 10.1007/s11055-020-01024-8
Berry R.B. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications, Version 2.2. / R.B. Berry, R. Brooks, C.E. Gamaldo et al. // Darien, Illinois: American Academy of Sleep Medicine, 2015. – 79 p.
Tallon-Baudry C. Induced gamma band activity during the delay of a visual short term memory task in humans / C. Tallon–Baudry, O. Bertrand, F. Peronnet et al. // Journal of neuroscience. – 1998. – Vol .18. – № 11. – P. 4244–4255. DOI: 10.1523/JNEUROSCI.18-11-04244.1998.
Yakovenko I.A. The study of rhythmic component coupling at the first stage of day sleep / I.A. Yakovenko, D.Y. Shumov, N.E. Petrenko et al. // Moscow University Biological Sciences Bulletin. – 2019. – Vol. 74. – № 2. – P. 98-102. DOI: 10.3103/S0096392519020111.
Peter-Derex L. Heterogeneity of arousals in human sleep: A stereo-electroencephalographic study / L. Peter-Derex, M. Magnin, H. Bastuji // Neuroimage. – 2015. – Vol. 123. – P.229–244. DOI: 10.1016/j.neuroimage.2015.07.057.
Ruby P. Dynamics of hippocampus and orbitofrontal cortex activity during arousing reactions from sleep: An intracranial electroencephalographic study / P. Ruby, M. Eskinazi, R. Bouet et al. // Human Brain Mapping. – 2021. – Vol .42. – № 16. – P. 5188-5203. DOI: 10.1002/hbm.25609. doi:10.1002/hbm.25609.
Cheremushkin E.A. Sleep and neurophysiological correlates of activation of consciousness on awakening / E.A. Cheremushkin, N.E. Petrenko, V.B. Dorokhov // Neuroscience and Behavioral Physiology. – 2022. – Vol. 52. – № 2. – P. 213–217. DOI: 10.1007/s11055-022-01226-2.