ОСОБЕННОСТИ ВОДНОГО РЕЖИМА ПОЙМЕННЫХ ЛУГОВ ВОДНО-БОЛОТНОГО УГОДЬЯ «ОЗЕРО КАРТАЛ»

Ассистент, Национальный университет гражданской защиты Украины
ОСОБЕННОСТИ ВОДНОГО РЕЖИМА ПОЙМЕННЫХ ЛУГОВ ВОДНО-БОЛОТНОГО УГОДЬЯ «ОЗЕРО КАРТАЛ»
Аннотация
В статье рассмотрено – особенности водного режима пойменных лугов, условия затопления поймы, схема формирования фитопродуктивности лугов в периоды разного затопления поймы.
Ключевые слова: пойма, пойменные луга, фитопродуктивность, обводненность.
Striuk T.Y.
Assistant, National University of Civil Protection of Ukraine
FEATURES WATER REGIME FLOODPLAIN MEADOWS WETLANDS "KARTAL LAKE"
Abstract
The article consideres especially the water regime of floodplain meadows, floodplain flooding conditions, the scheme forming phito productivity meadows in different periods of flooding floodplain.
Keywords: floodplain meadows, phito productivity, water cut.
Озеро Картал входит в систему западной группы Придунайских озер (Кагул, Картал, Кугурлуй-Ялпуг). Эти водоемы находятся на юге степной зоны Украины в пределах Ренийского района Одесской области в среднестепной подзоне Причерноморской среднестепной провинции Заднестровско-Причерноморской низинной области. Также озеро Картал является водно-болотным угодьем международного значения и входит в Рамсарский список.
К лугам относят растительные сообщества, основу которых составляют многолетние травянистые растения-мезофиты, требующие для своего развития умеренно влажные и умеренно богатые сравнительно теплые почвы с достаточной аэрацией. При увеличении влажности, а также при снижении почвенной температуры и содержания в почве кислорода луговые сообщества становятся болотами, где преобладают растения-гигрофиты. Резких границ между лугами и другими типами растительного покрова обычно не существует. К лугам относят также и ценозы с наличием галофитов при преобладании галомезофитов – солончаковые луга, – развивающиеся в условиях сухости климата при плохом дренаже, повышенном содержании в почвах легкорастворимых солей и формирующиеся на морских побережьях и террасах; рядом переходов они могут быть связаны с солонцами и солончаками.
Луга существуют в условиях сезонного климата, когда по сезонам года, а также от года к году изменяется поступление солнечной радиации и атмосферных осадков, а на ряде типов лугов, помимо того, при изменчивом гидрологическом режиме (колебание уровня почвенных и грунтовых вод, различие по годам в заливании полыми водами и др.). Это обуславливает динамику в обеспечении растений водой, элементами минерального питания и другими условиями, необходимыми для их произрастания. Изменяемость выраженности отдельных условий произрастания в течение года и от года к году имеют большое экологическое значение применительно к увлажнению [1, 2].
Луга, как и другие единицы растительности, представляют собой биогеоценозы. Луговой биоценоз – это комплекс групп живых организмов – высших и низших растений, грибов и различных представителей животного мира. В состав экотопа входит надземная среда (аэротоп) и косная часть почвы – эдафотоп, или почвенно-грунтовые условия. Основная особенность, отличающая биоценозы травянистых многолетних растений – луга или степи – от лесных, где определяющую ценотическую роль играют деревья и кустарники, это общая маломощность растительного яруса с ежегодным разрушением его по окончании вегетации растений. Поэтому факторы внешней среды – космические, атмосферные, гидрологические и эдафические – в большой степени определяют существование самих ценозов, обладающих относительно небольшой средообразующей способностью. Наиболее резкие влияния внешних условий, связанные с ценотической слабостью лугов, заключаются в сильном физическом воздействии на них выпадающих осадков, когда надземные части луговых ценозов могут оказаться полностью разрушенными при сильных ливневых дождях и граде. Весной и в начале лета луговые растения испытывают резкие перепады температур – от замерзания ночью до перегрева на солнце днем. При этом большинство луговых растений на разных этапах своего развития вынуждены функционировать в условиях полной солнечной радиации. Луговые ценозы в целом характеризуются слабым влиянием на компоненты природных экосистем. Однако на лугах в результате деятельности входящих в них организмов все же в какой-то степени создается специфический микроклимат. Так, внутри луговых травостоев по высотным горизонтам меняются световой и тепловой режимы, влажность воздуха и содержание в нем углекислого газа.
В связи с изменением условий произрастания в пределах ареалов отдельных видов, а также из-за сезонной и разногодичной динамичности среды в пределах любого лугового биоценоза большое значение имеет выносливость растений неблагоприятных условий – способность произрастать временно или постоянно в более или менее угнетенном состоянии в субоптимальных условиях по какому-либо из экологических факторов или по их комплексу. Это свойство приобретает высокое значение для растений, произрастающих в местах с резко изменяющимися эпизодически или периодически условиями произрастания, где у растений возникли приспособления к сохранению при наступлении неблагоприятного для роста периода определенной численности особей в угнетенном или покоящемся состоянии, чтобы после возврата «благоприятных условий» восстановить былое участие в фитоценозе. Степень устойчивости можно характеризовать с одной стороны изменением численности от оптимальных к неблагоприятным условиям произрастания, с другой – их реактивность, способность реагировать на улучшение условий произрастания. Растения испытывают на себе воздействие не единичных, изолированных в своем влиянии друг на друга экологических факторов, а их совокупности. Одна и та же выраженность какого-либо экологического фактора может оказать на растение различное влияние в зависимости от сочетания других факторов. По Л.Г. Раменскому [3], «одним из выражений совместного влияния факторов является их частичная взаимозаменяемость. В экологии нет неизменных нормативов, все отношения высоко подвижны». Все это вносит значительные трудности при классификации экологических факторов. Однако можно выделить три группы факторов:
- воздействующие через почву (водный режим почвы, состав почвенного воздуха, тепловой режим почвы);
- действующие на растения через их надземные органы (поступление солнечной радиации);
- связанные с воздействием человека (использование лугов как сенокосы, пастбища и для рекреации).
Обычно растения на лугах представлены не единичными экземплярами, относящимися к какому-либо виду, а совокупностью особей этого вида (ценотической популяцией). Каждая ценотическая популяция характеризуется числом растений, ее составляющих, и соотношением их возрастных групп. Возрастные группы соответствуют основным этапам жизни многолетних видов. Они представлены жизнеспособными семенами, находящимися в почве или на ее поверхности, еще не цветущими растениями, взрослыми особями, находящимися на вершине своего развития, цветущими и плодоносящими, и, наконец, растениями отмирающими, резко сокращающими рост, слабоцветущими или нецветущими. От характера возрастного строения популяций луговых растений, т. е. от соотношения в них возрастных групп, зависит как степень устойчивости луговых ценозов, так и возможности изменения их состава и строения. Возрастной спектр состава популяций дает возможность видам более полно использовать среду, выживать в критических или вообще неблагоприятных условиях.
Выделяется несколько групп местообитаний луговых ценозов, характеризующихся в основном двумя абиотическими факторами – степенью и характером увлажнения, а также богатством почвы. В прямой зависимости от них находятся состав и структура луговых ценозов, их динамические тенденции и продуктивность.
На суходолах хорошо различаются три группы местообитаний с соответствующим набором луговых сообществ: сухие луга (абсолютные суходолы), влажные (нормальные суходолы) и сырые (суходолы временно избыточного увлажнения). Низинные луга (всегда сырые) иногда делят на луга грунтового и ключевого питания; флористическая разница между ними ничтожна. На низинных лугах часто имеется примесь болотных растений и иногда бывает трудно провести четкие границы между болотистым лугом и болотом. Низинные луга грунтового питания приурочены к плоским низинам и котловинам с проточным увлажнением. Для них типичны богатые темноцветные дерновые почвы. Низинные луга ключевого питания, развивающиеся в местах выхода на поверхность движущихся грунтовых вод, по растительности не отличаются от предыдущего типа. Они занимают здесь периферические участки, окружающие топь или болото.
Пойменные, или заливные, луга, приуроченные к выработанным долинам рек, формируются в условиях периодического затопления вешними водами, образующимися в результате таяния снегового покрова, с одновременным отложением наилка. Характер растительности речных пойм зависит, прежде всего, от длительности затопления (так называемой поемности) и мощности наилка, причем оба эти фактора могут существенно изменяться год от года.
Поемность – важный экологический фактор отбора, влияющий на формообразование луговых видов и определяющий состав луговых пойменных ценозов. Поемность возрастает по направлению течения реки. В верховьях пойма заливается обычно на краткое время; в расширенных долинах низовий поемность может быть выражена в очень сильной степени. Реакция растений на затопление зависит от их состояния (покоя или активности) и длительности затопления. При затоплении в ранневесенний период, когда растения еще не тронулись в рост, они лучше выдерживают затопление, чем в период начала вегетации. Особенно неблагоприятно на луговые травы действует застой полых вод небольшой мощности в конце половодья при установлении теплой погоды, когда содержащийся в текучей воде кислород уже израсходован, а растения перешли в активное состояние и начали отрастать. Ежегодное отложение наилка – также важный экологический фактор, оказывающий непосредственное воздействие на виды растений, селящиеся в пойме. Перекрытие их наилком может надолго задержать начало вегетации или совсем погубить их. Наилок мощностью 1-2 см обычно не оказывают на луговые травы неблагоприятного влияния и даже хорошо сказываются на росте ценных в кормовом отношении видов. Однако увеличение мощности наилка, хотя бы на 1 см, отрицательно воздействует на их жизненное состояние. Чем больше у растений выражена способность к вегетативному размножению, тем лучше они выносят перекрытие наилком, так как они легко перемещают подземные органы с почками возобновления из более глубоких слоев к поверхности почвы. Погребение наилком, состоящим из рыхлых, несвязанных песчаных и супесчаных частиц, растения выносят лучше, чем суглинистым и глинистым, так как последний, подсыхая, образует плотную корку, через которую побеги могут пробиться лишь по трещинам. Поймы крупных рек представляют собой систему участков, различающихся по рельефу, механическому составу грунта, богатству почвы и увлажнению, сформировавшихся в результате расширения долины боковыми смещениями русла реки и отложения речных наносов – аллювия. Это обусловлено многими причинами, из которых, прежде всего, следует отметить гидрологический режим реки, уровень грунтовых вод, характер почвенного субстрата, растительный покров и климатические условия местности.
Продуктивность надземной массы пойм в основном зависит от комплекса гидрометеорологических условий. При этом гидрологическому режиму реки отводится ведущее значение, так как влияние этого режима чаще всего бывает специфичным (пойма затапливается талыми водами весной при таянии снега, иногда летом при обильных летних осадках).
Не рассматривая все гидрологические и метеорологические характеристики, имеющие значение для формирования фитопродуктивности естественных угодий, следует отметить, что из всего комплекса условий главным является режим затопления, характеризующийся продолжительностью, датами затопления, мощностью слоя воды, ее температурой, климатическими и метеорологическими условиями данного региона, типами почв, их засоление, рельефом. Такой комплекс формирует определенный ботанический состав пойменной растительности, ее урожайность, динамику роста и развития. От продолжительности паводкового затопления в весенне-летний период зависит не только общее влагонасыщение верхних горизонтов почвы в пойме, но и уровень залегания грунтовых вод, а следовательно, и уровень капиллярной каймы, что отражается на всех жизненных процессах растений, в том числе и на их продуктивности [4].
Пойменных лугов немного, но они дают больше половины сбора сена с естественных угодий и значительное количество подножного корма. Они продуктивнее и разнообразнее материковых лугов. Это хорошие пастбища, кроме того на них можно получить от 10-13 ц/га до 25-30 ц/га, иногда до 50-60 ц/га сена. Наиболее распространенными на территории ВБУ «Озеро Картал» являются тростниковые формации. Тростник обыкновенный обладает большой экологической пластичностью, он произрастает как на длительно затапливаемых участках, так и на участках с отсутствием ежегодного затопления. В условиях достаточной влагообеспеченности основным лимитирующим фактором является температурный режим. Как показывают наблюдения [4, 5], максимальный линейный прирост тростника наблюдается при средних суточных температурах воздуха 16-18 °С. После спада паводковых вод по мере понижения уровня грунтовых вод, а также в результате расходования влаги из верхнего корнеобитаемого слоя почвы роль метеорологических факторов в жизнедеятельности растений возрастает.
Длительное затопление (до первой декады июня) в условиях слабой проточной воды, прогревающейся в среднем за сутки выше 17 °С, ведет к снижению фитопродуктивности, деградации тростниковых лугов и замене тростника в травостое рогозом и другими малоценными в кормовом отношении видами. Сокращение продолжительности паводкового затопления ведет также к резкому снижению сена.
При определении фитопродуктивности пойменных лугов использована базовая модель оценки агроклиматических ресурсов формирования продуктивности сельскохозяйственных культур, предложенная А.Н. Полевым [6]. При этом допускается, что модель, применяемая по отношению к агроценозам используется при оценке фитопродуктивности луговых растений. Модель [7] описывает рост и развитие единичного растения, а биомасса посева определяется как произведение рассчитанной биомассы одного растения за каждые сутки на плотность посева. Модель учитывает некоторые эмпирические зависимости, охватывающие основные процессы жизнедеятельности растений, генетические особенности и физику среды обитания. Прирост биомассы определяется как разность между фотосинтезом и потерями на дыхание. Одним из регуляторов распределения прироста биомассы является относительная поверхность листа, которая рассчитывается в модели (отношение поверхности листа к его массе).
Процесс формирования фитомассы представляет собой совокупность многих физиологических процессов, интенсивность которых определяется биологическими особенностями растений, факторами окружающей среды, взаимосвязью между самими процессами.
Блок-схема модели формирования продуктивности лугов включает блоки основных физиологических процессов жизнедеятельности растений: фотосинтез, дыхание, рост, развитие, а также гидрометеорологический блок [7]. Хотя продуктивность посева определяется всем комплексом связей в системе почва-растение-атмосфера, при разработке модели использовались только те метеорологические и агрометеорологические данные о температуре и влажности воздуха, осадках, солнечной радиации, водно-физических свойств и влажности почвы, которые можно получить при помощи стандартных метеорологических и агрометеорологических наблюдений [6].
Базовая модель формирования продуктивности лугов имеет блочную структуру.
Блок фотосинтеза. Фотосинтез представляется формулой:
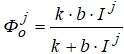 (1)
(1)
где ![]() - интенсивность фотосинтеза при определенных условиях влагообеспеченности в реальных условиях освещения, мгСО2/(дм2×г);
- интенсивность фотосинтеза при определенных условиях влагообеспеченности в реальных условиях освещения, мгСО2/(дм2×г);
k - интенсивность фотосинтеза при световом насыщении и нормальной концентрации СО2, мгСО2/(дм2×г);
b - начальный уклон световой кривой фотосинтеза, мгСО2/(дм2×г);
І - интенсивность фотосинтетически активной радиации (ФАР) внутри посева, кал/(см2×мин);
j - номер шага расчетного периода.
Для расчета фотосинтеза в онтогенезе в реальных условиях среды, которые отличаются от биологически оптимальных, используется выражение
![]() (2)
(2)
где ![]() - интенсивность фотосинтеза в реальных условиях среды, мгСО2/(дм2×г);
- интенсивность фотосинтеза в реальных условиях среды, мгСО2/(дм2×г);
![]() - онтогенетическая кривая фотосинтеза;
- онтогенетическая кривая фотосинтеза;
![]() - функции влияния факторов окружающей среды.
- функции влияния факторов окружающей среды.
Функции ![]() - нормированные и изменяются от 0 до 1.
- нормированные и изменяются от 0 до 1.
Суммарный фотосинтез посева за светлое время суток рассчитывается по формуле:
![]() (3)
(3)
где Ф - дневной фотосинтез посева на единицу площади, г/(м2×день);
ε =0,68 - коэффициент эффективности фотосинтеза;
L - площадь листьев м2/м2;
τ - продолжительность дня, час.
Блок дыхания. В отличие от процесса фотосинтеза способность к дыхательному газообмену имеют все органы растений. Потери на дыхание, связанное с поддержанием структурной организации тканей, и на дыхание, связанное с перемещением веществ, фотосинтезом и созданием новых структурных единиц, определяется по уравнению:
![]() (4)
(4)
где R - потери на дыхание, г/м2 ;
aR - онтогенетическая кривая дыхания;
c1 -коэффициент, который характеризует потери на поддержку структуры;
М - сухая биомасса посева, г/м2;
c2 - коэффициент, характеризующий потери, связанные с перемещением веществ, фотосинтезом и созданием новых структурных единиц;
j - порядковый номер декады расчета.
Блок роста. Прирост биомассы посева определяется остатком между суммарным фотосинтезом посева и потерями на дыхание:
ΔМ j = Ф j - R j (5)
Для описания роста отдельных органов растений используются ростовые уравнения в модифицированном виде, предложенные Ю.К. Росом [8].
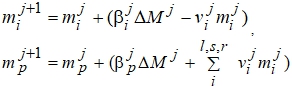 (6)
(6)
где mi – общая сухая биомасса отдельных органов iÎl,s,r,р (l – листья,
s - стебли, r - корни, p - репродуктивные органы), г/м2;
βi - функция перераспределения свежих "ассимилятов";
vi - функция перераспределения "старых" ассимилятов.
Увеличение площади листьев посева определяется при положительном приросте биомассы листьев по формуле:
![]() (7)
(7)
где z - удельная поверхность площади листьев, г/м2.
При отрицательном приросте биомассы листьев для описания роста ассимилирующей поверхности применяется соотношение:
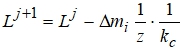 (8)
(8)
где kc=0,3 - параметр, характеризующий критическую величину уменьшения живой биомассы листьев, при которой начинается их отмирание.
ФАР, поглощенная посевом, рассчитывается по формуле:
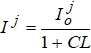 (9)
(9)
где ![]() - поглощенная солнечная радиация, кал/(см2/мин);
- поглощенная солнечная радиация, кал/(см2/мин);
С = 0,5 - эмпирическая постоянная величина;
L - площадь листьев, м2/м2.
Поток ФАР на верхнюю границу посева определяется во формуле:
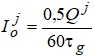 (10)
(10)
где Q - суммарная солнечная радиация, кал/ (см2/сут).
Суммарная солнечная радиация рассчитывается по формуле С.И. Сивкова [9]:
![]() (11)
(11)
где S - продолжительность солнечного сияния, час;
ho – полуденная высота Солнца.
Средняя за светлый период суток температура воздуха рассчитывается по формуле:
![]() (12)
(12)
где Тg , Тmax - соответственно средняя за день и максимальная температура воздуха;
ao, a1 - эмпирический коэффициент.
В соответствии с описанной структурой модели ее параметры разделены на четыре группы:
- параметры для расчета интенсивности фотосинтеза;
- параметры для расчета интенсивности дыхания;
- параметры для расчета динамики биомассы отдельных органов и всего растения, площади ассимилирующей поверхности;
- параметры, учитывающие значения коэффициентов уравнений регрессии для расчета средней за светлое время суток температуры воздуха.
В блок фотосинтеза входят параметры, характеризующие интенсивность протекания процесса фотосинтеза под влиянием факторов, которые непосредственно участвуют в самом процессе, а также те, которые отражают условия проведения процесса. Последние являются функциями влияния факторов среды на интенсивность процесса фотосинтеза.
Для роста тростниковых растений в условиях затопления необходимо учитывать интенсивность ФАР на поверхности воды и в ее толще, то есть необходимо определить ту глубину, при которой фотосинтез прекратится (рис. 1).
Точка компенсации (рис. 2) определялась в пределах интенсивности солнечной радиации от 0,02 до 0,03 кал·см2/мин и равна 0,026 кал·см2/мин. Таким образом, на глубине 1,25 м фотосинтез не сможет компенсировать потери на дыхание.
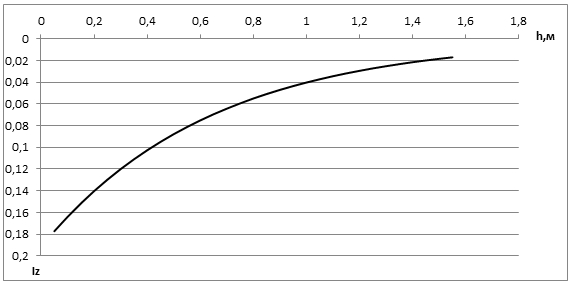
Рис. 1 - Ослабление солнечной радиации в толще воды
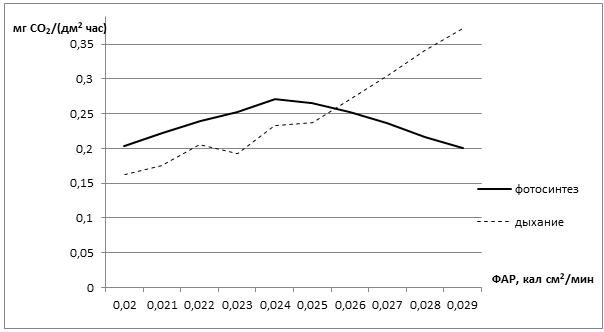
Рис. 2 - Точка компенсации
В работе использовались метеорологические данные по метеостанции Измаил. Данная модель позволила оценить продуктивность тростниковых лугов (рис. 3, 4) и разнотравных (рис. 5, 6) на сопредельных территориях озера Картал, при различных гидрологических условиях.
Для тростниковых лугов рассматривался период с марта по ноябрь.
Рис. 3 - Распределение прироста и массы тростниковых лугов при оптимальном увлажнении территории (весенние паводки)
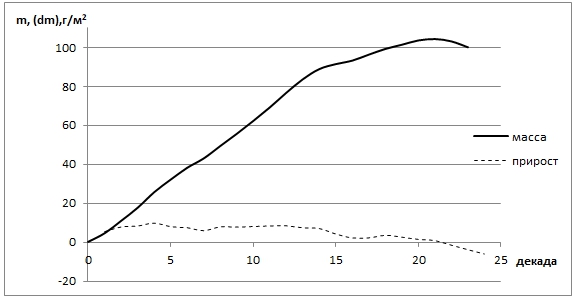
Рис. 4 - Распределение прироста и массы тростниковых лугов в условиях маловодного года
Как видно на рис. 3 рост фитомассы тростника идет на протяжении всего наблюдаемого периода, до конца июня наблюдается и прирост массы, который затем идет на спад. Фитопродуктивность тростника в условиях весеннего затопления составляет 813 г/м2 (сухого вещества). Таким образом, фитопродуктивность тростника на всей территории составит 81,3 ц/га.
Как видно из рис. 4 наблюдается рост фитомассы тростника с марта по середину июня, в период цветения тростника он становится более плавным. С середины апреля начинается волнообразный спад прироста массы. Спад фитомассы тростника начинается с концом плодоношения на 21 декаде (конец сентября). В условиях маловодного года, когда влагозарядка почвы полыми водами не происходит, наблюдается и снижение фитопродуктивности (10 ц/га).
Что касается разнотравных лугов, то в условиях весенне-летнего затопления, после спада воды, наблюдается резкий рост фитомассы (26,4 ц/га), но достигнув пика в конце мая отмечается спад прироста массы (рис. 5).
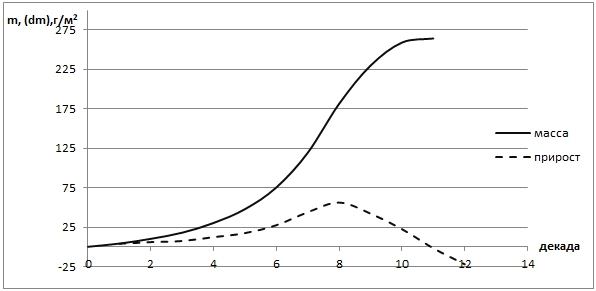
Рис. 5 - Распределение прироста и массы разнотравных лугов в условиях периодической обводненности территории
Для условий маловодного (засушливого) года наблюдается плавный рост фитомассы разнотравных лугов на всем протяжении наблюдаемого периода (рис. 6). По сравнению с предыдущим вариантом, наблюдается снижение фитопродуктивности в четыре раза.
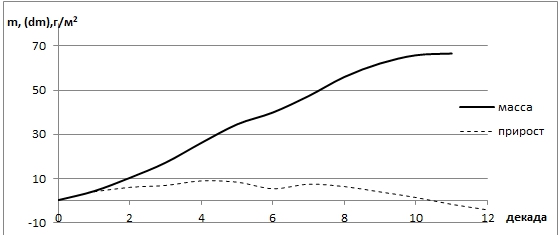
Рис. 6 - Распределение прироста и массы разнотравных лугов в условиях маловодного года
ВБУ «Озеро Картал» характеризуются разнообразием биотических сообществ, функционирование которых зависят от абиотических условий и степени их изменения под воздействием антропогенных факторов, которые негативно отразилось на биологическом разнообразии озера и прилегающей территории. Оптимизация водного (уровненного) режима будет способствовать сохранению и восстановлению биологического разнообразия и пойменных лугов, для которых периодическое затопление является важным фактором функционирования. При условии периодического затопления пойменных участков их фитопродуктивность значительно увеличится (81,3 ц/га и 26,4 ц/га), что позволяет использовать их ресурсы в качестве сенокосов и пастбищ.
Литература
- Бедарев С.А. Агрометеорология и лугопастбищное хозяйство / С.А. Бедарев. – Л.: Гидрометеоиздат, 1979. – 256 с.
- Бедарев С.А. К вопросу оценки и прогноза урожайности пойменных лугов на примере низовий р. Чу / С.А. Бедарев, Л.В. Лебедь // Тр. КазНИГМИ. – 1974. – вып. 47. – С. 72-75.
- Раменский Л.Г. Классификация земель по их растительному покрову / Л.Г. Раменский // Пробл. Ботаники. – 1950. – вып.1 – 502 с.
- Лебедь Л.В. Агрометеорологические условия развития пойменной растительности / Л.В. Лебедь // Тр. КазНИГМИ. – 1974. – вып.47. – С. 66-71.
- Раменский Л.Г. Введение в комплексное почвенно-геоботаническое исследование земель / Л.Г. Раменский. – М. – 1938. – 648 с.
- Польовий А.М. Моделювання гідрометеорологічного режиму та продуктивності агроекосистем: Навчальний посібник / А.М. Польовий. – К.: КНТ, 2007. – 344 с.
- Тооминг Х.Г. Солнечная радиация и формирование урожая / Х.Г. Тооминг. – Л.: Гидрометеоиздат, 1977. – 200 с.
- Росс Ю.К. Радиационный режим и агротектоника растительного покрова / Ю.К. Росс. – Л.: Гидрометеоиздат, 1975. – 314 с.
- Сивков С.И. Методы расчета характеристик солнечной радиации/ С.И. Сивков. – Л.: Гидрометеоиздат, 1961. – 227 с.