МИКРОСКОПИЧЕСКОЕ СТРОЕНИЕ ПОЛОВЫХ ЖЕЛЕЗ ДОМАШНЕГО ЯПОНСКОГО ПЕРЕПЕЛА

Савельева А.Ю.
ORCID: 0000-0001-7186-3039, Кандидат ветеринарных наук, Красноярский государственный аграрный университет
МИКРОСКОПИЧЕСКОЕ СТРОЕНИЕ ПОЛОВЫХ ЖЕЛЕЗ ДОМАШНЕГО ЯПОНСКОГО ПЕРЕПЕЛА
Аннотация
Выявлено, что относительная масса обоих гонад самца половозрелого японского перепела составляет 1,95%, правая гонада весит больше и имеет и более вытянутую форму относительно левой. Установлены морфологические особенности всех клеточных форм сперматогенного эпителия. Выявлено, что нет четкого морфологического отличия между сперматогониями и растущими сперматоцитами.
Ключевые слова: японский перепел, семенники, микроструктура, извитые канальцы семенника, сперматогенный эпителий, клетки Сертоли
Savelyeva A.J.
ORCID: 0000-0001-7186-3039, PhD in Veterinary Medicine and Science, Krasnoyarsk State Agrarian University
MICROSCOPIC STRUCTURE OF GONADS DOMESTIC JAPANESE QUAIL
Abstract
Studies have revealed that sexually mature quаil testicles’ mass is 1,95% of his total body mass. The right gonad is bigger heavier and more elongated then the left one. It was found morphological features of all cellular forms of spermatogenic epithelium. It was revealed that there is no clear morphological differences between the spermatogonia and growing spermatocytes.
Keywords: Japanese quail, testes, convoluted tubules, microstructure, spermatogenic epithelium, Sertoli cells
Введение. Вопросу репродуктивной системы самок сельскохозяйственной птицы, как объекта экономически выгодного для сельского хозяйства, привлекательного для науки, ценного для пищевой промышленности посвящено большое количество научных работ отечественных и зарубежных морфологов. Достаточно глубоко исследованы половые органы кур, индеек, уток, перепелов [1], в то же время вопросы морфологии половой системы самцов домашней птицы раскрыты недостаточно, имеющиеся сведения весьма разрозненные и касаются в основном анатомии.
Материалы и методы исследования. Объектом исследования послужили самцы 60-суточного возраста. Материалом для исследования послужили 12 семенников от шести голов.
Морфометрия. Взвешивание органов производили на весах аналитических Сартогосм «СЕ 224-С», промеры производили при помощи штангенциркуля с простой регулировкой.
Гистологические методы. Фиксацию материала проводили в 10% нейтральном растворе формалина, обезвоживание проводили по методу М.О. Зайцева [2]. Уплотняли материал в парафине при температуре 60°С. Поперечные и продольные срезы толщиной 6-7 мкм изготавливали на санном микротоме МС-2. Для получения обзорных препаратов гистологические срезы окрашивали гематоксилином Эрлиха и эозином [3].
Микроскопия и микроморфометрия. Изучение и микрофотографирование гистологических препаратов проводили под бинокулярным световым микроскопом MS (Austria) при увеличении в 100, 400, 1000 раз. Структурную микроморфометрию осуществляли с помощью окуляр-микрометра МОВ-1-15х (ГОСТ 15150-69). Оценку достоверности различий проводили методом вариационной статистики с использованием t-критерия Стъюдента. Различия считали значимыми, если вероятность случайности не превышала 5% (Р<0,05).
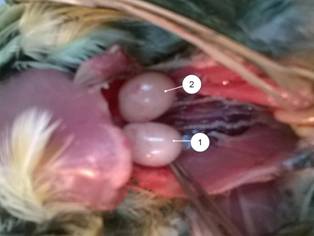
Результаты и обсуждение. Живая масса самцов в исследуемом возрасте составляет 165,24±2,81 гр, относительная масса обоих семенников равна 1,95%. Семенники имеют яйцевидную или слегка бобовидную форму, располагаются в передне-поясничной области под пояснично-крестцовой костью. В отличие от самок, у самцов японского перепела развиты обе гонады, правый семенник слегка более вытянут (рис. 1), левый более компактный и в 1,15 раза легче (Р < 0,001) (табл. 1).
Таблица 1 – Линейные размеры и масса гонад
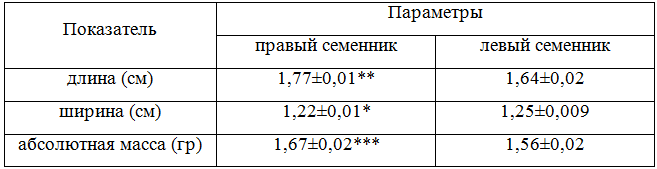
Примечание: (М±m, n=6), достоверность различий правого семенника по отношению к левому, где* – Р < 0,05;** – Р < 0,01; *** – Р < 0,001.
Снаружи гонада покрыта белочной капсулой из плотной оформленной соединительной ткани, толщина ее колеблется в разных участках от 19,6 до 58,8 мкм. Капсула пронизана кровеносными сосудами. Элементы волокнистой стромы погружаются вглубь паренхимы, также сопровождаемые сосудами.
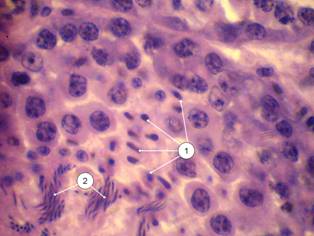
Микроскопическое строение паренхимы соответствует таковому у представителей класса млекопитающих. На поперечном срезе паренхима представлена многочисленными срезами извитых канальцев и интерстициальной тканью. Диаметр канальцев составляет 230±12,7 мкм. Соединительнотканная стенка канальца толщиной около 2,7 мкм отграничивает популяции сперматогенного эпителия, на его долю приходится около 66,3±4,92 мкм просвета канальца. Эпителий представлен поддерживающими клетками, или сустеноцитами, и четырьмя разновидностями половых клеток, находящихся на последовательных стадиях сперматогенеза (рис. 2).
 |
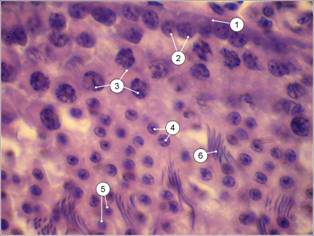 |
| Рис. 1. – Семенники домашнего японского перепела 60-суточного возраста (печень, желудок, кишечник изъяты): 1 – правый, 2 – левый | Рис. 2 – Сперматогенный эпителий (гематоксилин Эрлиха-эозин) об. 100х, ок. 10х: 1 – оболочка канальца; 2 – сперматогонии; 3 – сперматоциты I порядка; 4 – сперматоциты II порядка; 5 – сперматиды; 6 – спермии |
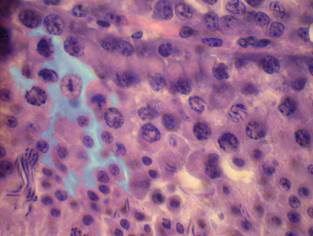
Ядра сустеноцитов расположены ближе к периферии извитого канальца, но не под самой его оболочкой, как это часто наблюдается у млекопитающих. Ядро крупное, неправильное овальное, диаметр составляет 7,61±1,81 мкм, в нём просматриваются мелкие зерна хроматина и обычно не более одного ядрышка, кариолимфа хромофобная, что делает сустеноциты хорошо заметными среди остальной клеточной массы, заполняющей каналец. Латеральные отростки и впячивания плазмолеммы при данном уровне микроскопии просматриваются нечетко, мы обработали фотографию в графическом редакторе для выявления клеточных границ (рис. 3).
 |
 |
| Рис. 3 – Сустеноцит (гематоксилин Эрлиха-эозин) об. 100х, ок. 10х: цитоплазматические отростки выделены голубым цветом | Рис. 4 – Стадия формирования в извитых канальцах: (гематоксилин Эрлиха-эозин) об. 100х, ок. 10х: 1 – начальная стадия превращений сперматид в спермии (изменение формы ядра, уменьшение объема цитоплазмы); 2 – колонии спермиев |
Сперматогенные клетки развиваются в углублениях цитоплазмы сустеноцитов, образуя необособленные колонии. Сперматогонии – округлые клетки, лежат на самой периферии канальца в один ряд, выделяются хромофильными ядрами. Растущие сперматоциты первого порядка расположены глубже и размеры их ядер крупнее предыдущей стадии в 1,15 раза (Р<0,01). Ядра клеток в связи с диплоидностью также, как и ядра первых, ярко окрашены. Сперматоциты II порядка и сперматиды либо лежат тяжами по 5-12 клеток, либо хаотично. Диаметр гораздо более светлых и менее плоидных ядер сперматоцитов II порядка меньше диаметра сперматогоний и сперматоцитов I порядка в 1,2 и в 1,4 раза (Р<0,05) соответственно. Диаметр ядер сперматид в 1,6 меньше диаметра сперматогоний (Р<0,001), в 1,9 – растущих сперматоцитов (Р<0,001), в 1,3 – сперматоцитов II порядка (табл. 2).
Таблица 2 – Диаметр половых клеток сперматогенного эпителия
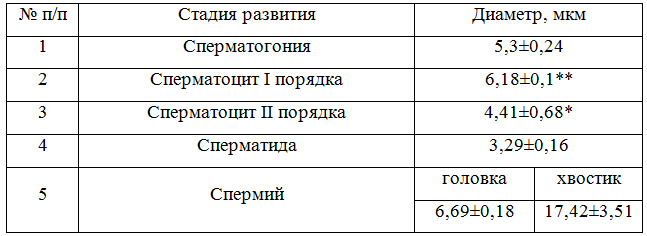
Примечание: (М±m n=10), достоверность различий каждой стадии развития половой клетки по отношению к предыдущей, где * – Р < 0,05;** – Р < 0,01; *** – Р < 0,001.
Стадия формирования представлена переходными этапами преобразования ядра из округлой в вытянутую форму, при этом заметно уменьшение объема не только ядра, но и цитоплазмы (рис. 4). Сформированные спермии сосредоточены в центральной части канальца, хотя палочковидная головка может внедряться в периферические слои эпителия. Нередко встречаются небольшие группки спермиев, головки которых расположены параллельными пучками, соотношение составляет длины головки и хвостика составляет 1:2,6.
Выводы
- По форме и по массе левая и правая гонады домашнего японского перепела неравнозначны: правая гонада весит больше и имеет и более вытянутую форму.
- Микроскопически паренхима семенника содержит все типы клеток сперматогенного эпителия, а также поддерживающие клетки и интерстициальную ткань.
- Нет четко выраженного различия в размерах сперматогоний и растущих сперматоцитов, что нехарактерно для млекопитающих.
Литература
- Савельева, А.Ю. Морфология репродуктивных органов перепелок: монография / А.Ю. Савельева; Краснояр. гос. аграр. ун-т. – Красноярск, 2011. – 129 с.
- Зайцев, М.О. «Новая» проводка гистологических тканей [Электронный ресурс]. URL: http://www.evrika.ru/show/283 (дата обращения 14.10.2015).
- Семченко, В.В. Гистологическая техника / В.В. Семченко, С.А Барашкова, В.Н. Артемьев. – Омск, 2003. – 152 с.
References
- Savel'eva, A.Ju. Morfologija reproduktivnyh organov perepelok: monografija / A.Ju. Savel'eva; Krasnojar. gos. agrar. un-t. – Krasnojarsk, 2011. – 129 s.
- Zajcev, M.O. «Novaja» provodka gistologicheskih tkanej [Jelektronnyj resurs]. URL: http://www.evrika.ru/show/283 (data obrashhenija 10.2015).
- Semchenko, V.V. Gistologicheskaja tehnika / V.V. Semchenko, S.A Barashkova, V.N. Artem'ev. – Omsk, 2003. – 152 s.