ПЛАСТИЧНОСТЬ СИНАПСОВ УЛИТКИ В СУБСЕКУНДНОМ ВРЕМЕННОМ ИНТЕРВАЛЕ

ПЛАСТИЧНОСТЬ СИНАПСОВ УЛИТКИ В СУБСЕКУНДНОМ ВРЕМЕННОМ ИНТЕРВАЛЕ
Научная статья
Палихова Т.А.1, *, Соколов Е.Н.2, Черноризов А.М.3
1 ORCID: 0000-0002-1170-0438,
1, 2, 3 Кафедра психофизиологии факультета психологии Московского Государственного Университета им. М.В. Ломоносова, Москва, Россия
* Корреспондирующий автор (palikhova[at]mail.ru)
АннотацияМозг отвечает на единичные сенсорные сигналы во временных интервалах менее одной секунды. В этот период регистрируются суммарные вызванные потенциалы и ответы отдельных нейронов, когда происходят изменения связей между нейронами. Пластичность во время ответа на единичный сенсорный стимул получила название немедленной пластичности [11]. Данная статья посвящена исследованию немедленной синаптической пластичности на идентифицирумых синапсах виноградной улитки методом одновременной внутриклеточной регистрации активности пре- и постсинаптических идентифицированных нейронов.
Ключевые слова: виноградная улитка, сенсорные нейроны, командные нейроны, идентифицированный синапс, немедленная пластичность.
SUBSECOND TIME-DEPENDENT PLASTICITY IN SNAIL,S SYNAPSES
Research article
Palikhova T.A.1, *, Sokolov E.N.2, Chernorizov A.M.3
1 ORCID: 0000-0002-1170-0438,
1, 2, 3 Department of Psychophysiology, Faculty of Psychology, Lomonosov Moscow State University, Moscow, Russia
* Corresponding author (palikhova[at]mail.ru)
AbstractResponse of the brain to the single sensory stimuli lasts about second. This period compound potentials and responses of neurons might be recorded, when connections within brain are changing. The change during response to a single stimulus has been named immediate plasticity [11]. The paper presented devotes to the investigations of immediate plasticity by simultaneous intracellular recording of electrical activity synaptically connected identified neurons of the snails Helix pomatia and Helix lucorum.
Keywords: snail, sensory neurons, command neurons, identified synapses, immediate plasticity.
ВступлениеСтатья посвящена вопросу соотношения восприятия и памяти во время ответа мозга и его элементов на одиночный сенсорный стимул. Е.Н. Соколов предложил для этого процесса термин «немедленная пластичность» и были проведены эксперименты для ее изучения. Был подготовлен текст статьи, но не был опубликован в связи с уходом Е.Н. Соколова в 2008 году. Сегодня мы, его ученики и коллеги в преддверии 100-летнего юбилея Евгения Николаевича Соколова в 2000 году подготовили не изданные при его жизни работы, которые дополнят созданную им психофизиологическую концепцию «Человек – Нейрон – Модель».
Introduction
A change of synaptic efficiency during a response to a single sensory stimulus, so called immediate plasticity [11] differs from short-term plasticity or long-term plasticity widely well-known as a mechanism for learning and memory. To study immediate plasticity in details we used paired simultaneous recordings from the identified presynaptic sensory neurons (SN) and postsynaptic command neurons (CN) of the parietal ganglia of a land snails Helix pomatia L. and Helix lucoruim L. The SNs have local receptive fields at surface of viscera [9], and the giant parietal neurons are known as the CN of the snail’s avoidance behavior [2]. Identified inputs from the identified SNs to the parietal CNs are cholinergic [6] and have complex morphological structure (Arakelov et al., 1991). The main experimental task of the research was to study the dynamics of elementary (unitary) excitatory postsynaptic potentials (eEPSPs) and compound excitatory postsynaptic potentials (cEPSPs = a sum of eEPSPs) recorded from a CN in response to single spikes elicited in the presynaptic SN by sensory stimulation or by intracellular current injection.. It was shown that the synapses between SN and CN may display depression and/or potentiation on sub-second time scale in dependence on presynaptic spike number and frequency pattern during a response. The results allow to suggest that sub-second timed synaptic plasticity may really participate in the formation of cEPSPs’ late components of cEPSPs and in associative plasticity of the responses of the CN to the sensory stimuli.
Material and Methods
Preparations
Experiments were performed on preparations of adult land snails Helix lucorum and Helix pomatia. The experiments were conducted in accordance with International legislation on the use of experimental animals. The researchers were guided by recommendations of the code of ethics adopted by the Council for International Organizations of Medical Sciences, by Russian legislation regulating biomedical research on animals and Bioethics Commission at Lomonosov MSU. Low-temperature (8-12oC) anesthesia was used during operations [6]. The preparation consisting of central ganglia, mantle collar and viscera was kept in a dish with a standard physiological solution for cold-blooded animals (Sokolov, Palikhova, 1999b).
RecordingIntracellular, somatic whole-cell current-clamp recordings were obtained simultaneously from the identified presynaptic SN and postsynaptic CN. Identification of the CNs and the SNs was based on the morphological and physiological criteria [1]. Intracellular microelectrodes filled with 2.5 M KCl had resistance 10 - 20 Mom for pre- and 1-5 Mom for postsynaptic neurons. Large soma size of CN allowed using low-resistance pipettes to record low-amplitude postsynaptic potentials. Signals obtained from SN and CN were amplified with MEZ-8201 (Nihon Kohden, Japan) and MS-03 (Minsk, Belorussia) amplifiers. Date were stored on computer by the analog to digital converter and program DS (Digiscope, Moscow) for monitoring, recording and subsequent analysis.
Stimulation
Intracellular current injection and tactile stimulation of the visceral organs surface were used to elicit spikes in the sensory neurons. Currents were injected into a cell through the recording microelectrode by means of a bridge circuit. Temporal parameters of the injected current depended on the experimental task. For rhythmic stimulation within a range of frequencies from 0,005 to 10 Hz short (5-50 ms) current pulses (1 - 10 nA) were used. Direct current injections into the SN had been used to trigger trains of presynaptic spikes that simulated trains of spikes elicited by sensory stimuli. Calibrated hairs (0.1 - 0.2 mm, 0.4 - 1.5 G) connected with a solenoid and operated by an electro-stimulator MSE-3R (Nihon kohden, Japan) were used for tactile stimulation. For electric stimulation of the skin surface paired silver electrodes with tip diameter 1 mm were used. Data from presynaptic sensory LPa7 and LPa9 and postsynaptic command neurons LPa3 and RPa3 were studied systematically in about 50 animals.
Results
Identified SN, CN and synapses
Cells’ somas of identified CN of snail’s avoidance behavior are located at the rostral part of the parietal ganglia and have size about 200 mkm in diameter. The parietal sensory neurons are small cells (about 20 mkm in diameter of soma) in the middle part of the left parietal ganglia. Responses of the SNs recorded to mechanic or electric stimulation of local areas within surface of viscera were presented by several action potentials (AP) generated without any depolarization shift in membrane potential of the soma (Fig. 1A). Each AP generated by the SN was accompanied by an eEPSP in CN (Fig. 1A). Brief mechanic or electric stimulation within the receptive field evoked in the CN a cEPSP as a sum of eEPSPs (see figure 1A). Thus, brief local tactile or electrical stimulation of the SNs receptive field elicits a burst of spikes in the presynaptic SN and eEPSP /cEPSP in the parietal CN.
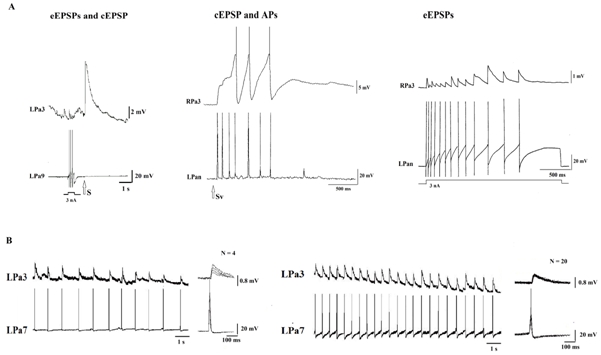
Fig. 1 – Amplitudes of eEPSPs in CN are changed during patterned train of presynaptic spikes in SN
Trains of APs elicited in an identifiable SN by brief sensory stimuli were consisted of 5 - 10 spikes. Duration of a spike train was as large as the duration of the cEPSPs recorded simultaneously in a CN (see fig. 1A). Interspike intervals within the trains varied in range from 50 ms at the beginning to 500 ms - 1 s at the end. In several experiments there it was possible to isolate eEPSPs from cEPSPs elicited by the sensory stimuli. The experiments have shown that the amplitude of those eEPSPs decreased at the beginning and increased at the tail of the compound responses of the CN. To simulate the results of a local sensory stimulation, trains of APs were triggered in a SN by depolarizing current injection. The simplest train was a sequence of two APs. If intervals between two neighboring APs were in the range of 50 – 200 ms, the eEPSP to the second AP was significantly reduced demonstrating immediate depression (see fig . 1A). If, however, the doubled APs were followed after 500 ms - 10 s with another AP, the eEPSP amplitude evoked by this AP was enhanced with respect to the initial value suggesting immediate potentiation (see fig 1A). Increasing in the number and frequency of APs in beginning of a train evokes a more pronounced potentiation and prolongs its effect. For example, a train of ten presynaptic APs evoked an increase of the eEPSPs’ amplitudes that could be revealed several minutes after the high-frequency stimulation. The potentiation in both cases was identified as posttetanic potentiation.
Synaptic responses during rhythmic stimulation
Rhythmic sensory stimulation results in habituation of the cEPSP in a CN within wide spectrum of frequencies and intensities of stimulation. Habituated (decreased) amplitudes of cEPSPs might be recovered by high frequency stimulation of another sensory input. Rhythmic spike generation in the presynaptic SN results in changes of amplitudes of a CN eEPSP in frequency-dependent mode. Magnitudes of eEPSPs are significantly decreased under high (2 Hz or more) and low (1 Hz or less) frequencies of presynaptic APs (see fig. 1B, left). Low frequency depression is very similar to depression shown for cEPSPs in the snails CN [10]. It’s interesting that there some middle meanings of the frequencies exist (1 Hz - 2 Hz), at which the amplitudes of eEPSPs don’t change (see fig. 1B, on the right).
Discussion
Brief local tactile or electrical stimulation elicits a burst of spikes in the SNs and eEPSP / cEPSP in the parietal CNs. Frequency of spikes is decreased within the burst from high values (10 - 20 Hz) at the beginning to low ones (1 - 2 Hz) at the tail of the AP burst. Simulation of the spike burst by intracellular current injection into a SN results in changes of eEPSP amplitudes recorded from a CN. The amplitude of eEPSPs is decreased at the beginning of the burst and increased when the frequency of spikes was reduced to 2 Hz. Amplitudes of eEPSPs evoked by low frequency spikes might be higher than the amplitude of eEPSP evoked by first spike in the burst. However a prolongation of low frequency spikes generation results in a secondary decreasing of eEPSP amplitudes. These data may be used to explain often registered two components of cEPSP (‘two-humped’) in CNs as a result of nonlinear dynamics of eEPSPs’ magnitudes during presynaptic burst of spikes on sub-second time scale.
Apparently, the nonlinear dynamics of eEPSPs is a result of interplay between high frequency depression, posttetanic potentiation and low frequency depression of the synapses between SNs and CNs. So, well-known different habituation dynamics of cEPSPs in dependence on frequency and intensity of sensory stimuli can be explained by interplay between high frequency depression and posttetanic potentiation of eEPSPs. Elevation of presynaptic Ca2+ concentration evoked by burst of spikes during response to the sensory stimulus can explain also selectivity of associative synaptic learning by influence of elevated Ca2+ to the presynaptic receptors. Sensitization of presynaptic receptors for the back-messengers (like neurotrophins or NO) after preliminary sensory evoked spike burst may be interpreted as a base for a “presynaptic waiting hypothesis” [7].
We can assume the habituation of synaptic transmission in the identified synapses of Helix is a summation of opposite directed plastic changes - depression and potentiation. One might predict existence of some frequencies at which processes of depression and potentiation compensate each other. Our experimental data confirm the supposition by demonstrating that there some frequencies exist (1 Hz - 2 Hz), at which the amplitudes of eEPSPs aren’t changed (see fig. 1 B, on the right).
Conclusion
It was shown that the synapses between SN and CN may display depression and/or potentiation already on sub-second time scale in dependence on presynaptic spike number and frequency dynamics (frequency pattern) during a response. The results allow suggesting that such sub-second timed immediate synaptic plasticity may really affect associative plasticity of the responses of the CN to the sensory stimuli. The data of the research extend the influence of models of time-dependent synaptic plasticity of Hebbian and/or Pavlovian types [3, 4] from the field of supra-second scales to the area of sub-second time intervals.
| Конфликт интересов Не указан. | Conflict of Interest None declared. |
Список литературы / References
- Arakelov G .G. Structural and functional analysis of monosynaptic connections between identified neurons of Helix lucorum /D.A. Sakharov and W. Winlow / Arakelov G .G., Marakujeva I.V., Palikhova T.A. //Simpler nervous systems. Manchester University Press.. Manchester. UK. – 1991. - P. 258-269.
- Balaban P.M. A system of command neurons in snail's escape behavior. Acta Neurobiol. Exp. – 1979. - № – P. 97-107.
- Butts D.A.The applicability of spike time dependent plasticity to development. Front. Synaptic Neuroscience / Butts D.A., Kanold P.O.. - 2010. - №2 1-9. doi: 10.3389/fnsyn.2010.00030.
- Caporale N. Spike Timing-Dependent Plasticity: A Hebbian Learning Rule. Rev. Neurosci / Caporale N., Dan. J.. – 2008 - № 31. – P. 25 - 46.
- Ierusalimski V.N. Nervous system and neural maps in Gastropod Helix lucorum L. Neurosci / Ierusalimski V.N., Zakharov I.S., Palikhova T.A.,. Comp. Physiol. – 1994. - № – P. 13-22.
- Kerkut G.A. The use of snail neurons in neurophysiological studies. In: The Missing Pieces. University of Southampton. Southampton. UK. – 1969.
- Palikhova, TA. Presynaptic waiting of a response: a hypothesis based on snail's identifiable synapses data [Proceeding of the13th Neuropharmacology Conference. “Post Translational Modifications of Protein Structure and Synaptic Function”. Official Satellite to the 2003 Meeting of the Society for Neuroscience] New Orleans, USA. - 2003. - November 5-7.
- Palikhova T.A. Cholinergic sensory inputs to command neurons in edible snail. Bulletin of Experimental Biology and Medicine. Springer New York Consultants Bureau / Palikhova T.A., Abramova M.S., Pivovarov A.S.. 2006142, 275-278.
- Palikhova, T. A.(1992). Monosynaptic connections in the central nervous system of the edible snail: receptive fields of presynaptic neurons / Palikhova, TA, Arakelov, GG.. Neurosci. Behav. Physiol. 22, 14-16.
- Shekhter E.D. Receptive field of command neuron / E.N. Sokolov and L.A. Shmelev . / / Neurocybernetic analysis of the mechanisms of behavior. Russia, Moscow: – 1985. - P. 64-84. [Шехтер Е.Д., Аракелов Г.Г. Рецептивное поле командного нейрона Соколов Е.Н., Шмелев Л.А. (ред.) Нейрокибернетический анализ механизмов поведения. М.: Наука. - 1985. – С. 64-84.]
- Sokolov E.N. Immediate plasticity of identifiable synapses in the land snails Helix lucorum / Sokolov E.N., Palikhova T.A. Acta Neurobiol. Exp. -1999a. - № P. 161-169.
- Sokolov E.N. Elementary and compound postsynaptic potentials in the defensive command neurons of Helix lucorum / Sokolov E.N., Palikhova T.A.. Acta Biologica Hungarica. - 1999b. - №50. – P. 1-11.