СОЧЕТАННЫЕ ИЗМЕНЕНИЯ ЭКСПРЕССИИ ГЕНОВ CSTB И ACAP3 ПРИ СИМПТОМАТИЧЕСКОЙ ЭПИЛЕПСИИ И БОЛЕЗНИ ПАРКИНСОНА

Белоцерковская Е.В.1, Сучкова И.О.2, Борисова Е.В.3, Боровкова Н.К.4, Павлинова Л.И.5, Паткин Е.Л.6
1ORCID: 0000-0003-3985-9552, кандидат биологических наук, научный сотрудник, 2ORCID: 0000-0003-2127-0459, кандидат биологических наук, старший научный сотрудник, 3ORCID: 0000-0002-7057-3096, кандидат медицинских наук, зав. отделением, 4ORCID: 0000-0003-1804-6368, аспирант, 5ORCID: 0000-0002-5848-5544, кандидат биологических наук, старший научный сотрудник,ФГБУН Институт физиологии им. И.П. Павлова, ведущий научный сотрудник, 6ORCID: 0000-0002-6292-4167, Доктор биологических наук, профессор,
ФГБНУ «Институт экспериментальной медицины»
Работа поддержана грантом РФФИ № 15-04-04642-a
СОЧЕТАННЫЕ ИЗМЕНЕНИЯ ЭКСПРЕССИИ ГЕНОВ CSTB И ACAP3 ПРИ СИМПТОМАТИЧЕСКОЙ ЭПИЛЕПСИИ И БОЛЕЗНИ ПАРКИНСОНА
Аннотация
С помощью ПЦР в режиме реального времени проведен анализ уровня мРНК генов центаурина бета 5 (ACAP3) и цистатина Б (CSTB) в лейкоцитах периферической крови у здоровых людей, а также у пациентов с симптоматической эпилепсией и болезнью Паркинсона (форма с ранним дебютом). По сравнению с контролем обнаружено повышение уровня мРНК гена ACAP3 у мужчин и женщин с эпилепсией (в 2,5- и 5-раз соответственно), а также у женщин с болезнью Паркинсона (примерно в 6 раз). Кроме того, у женщин наблюдалось снижение уровня мРНК гена CSTB при эпилепсии (примерно в 3 раза) и при болезни Паркинсона (примерно в 2.5 раза). Полученные данные указывают на возможное участие исследованных генов в патогенезе симптоматической эпилепсии и болезни Паркинсона.
Ключевые слова: периферическая венозная кровь, ПЦР в режиме реального времени, экспрессия генов ACAP3 и CSTB gene expression, симптоматическая эпилепсия, болезнь Паркинсона.
Belotserkovskaya E.V.1, Suchkova I.O.2, Borisova E.V.3, Borovkova N.K.4, Pavlinova L.I.5, Patkin E.L.6
1ORCID: 0000-0003-3985-9552, PhD in Biology, scientist, 2ORCID: 0000-0003-2127-0459, PhD in Biology, 3ORCID: 0000-0002-7057-3096, MD, The Neurology, 4 ORCID: 0000-0003-1804-6368, Postgraduate student, 5ORCID: 0000-0002-5848-5544, PhD in Biology, leading scientist, 6ORCID: 0000-0002-6292-4167, PhD in Biology, Professor,
FSBSI "The institute of experimental medicine".
The work was supported by RFBR grant № 15-04-04642-а
CONCURRENT CHANGES OF CSTB AND ACAP3 GENES EXPRESSION IN SYMPTOMATIC EPILEPSY AND PARKINSON’S DISEASE
Abstract
Real-time PCR analysis of the mRNA level of the genes of cetaurin beta 5 (ACAP3) and cystatin B (CSTB) in peripheral blood leukocytes in healthy people and in patients with symptomatic epilepsy and Parkinson's disease (early debut form) was performed. Compared to the control, an increase in the mRNA level of the ACAP3 gene in men and women with epilepsy (2.5 and 5 times, respectively) was found, as well as in women with Parkinson's disease (approximately 6-fold). In addition, a decrease in the mRNA level of the CSTB gene was observed in women with epilepsy (approximately 3-fold) and Parkinson's disease (approximately 2.5-fold). The data obtained indicate the possible involvement of the investigated genes in the pathogenesis of symptomatic epilepsy and Parkinson's disease.
Keywords: periphery venous blood, real-time PCR, ACAP3 and CSTB gene expression, symptomatic epilepsy, Parkinson’s Disease.
Introduction
Epilepsy is one of the most common neurological disorders with neurodegenerative features connected with numerous changes at the cellular or synaptic level such as defective arborization of the neurons and synaptic reorganization [1, P. 99]. Parkinson’s disease (PD) is a neurodegenerative disorder affecting both dopaminergic and non-dopaminergic neuronal systems. Pathogenesis of PD is associated with the destruction of specific neuron populations, synaptic damage and the presence of misfolded or aggregated proteins [2, P. 151], [3, P. 12]. The clinical findings support the assumption of the neurological comorbidity of epilepsy with PD [4, P. 45], [5, P. 1613], [6, P. 349]. The link between neurodegenerative diseases (including PD) and progressive myoclonus epilepsies [7, P. 390], [8, P. 293], [9, P. 61] may be explained by some common molecular and cellular mechanisms due to dysfunction of the mutual genes responsible for the structure and functioning of nerve cells.
Cystatin B protein (also known as CSTB; StB; stefin B) is one of numerous proteins possibly involved in process of neurodegeneration. Cystatin B is a member of a large family of intracellular inhibitors of cysteine proteinases [10, P. 5406]. It downregulates the cathepsins and interacts with a number of proteins involving in cell growth and cytoskeletal organization as part of a multiprotein complex [11, P. 2941], [12, P. 557], [13, P. 205]. It is well established that a cause of progressive myoclonus epilepsy (EPM1) of Unverricht-Lundborg type is mutations in the CSTB gene [14, P. 847], [15, P. 1731], [16, P. 5910]. The cystatin B deficiency leads to oxidative stress and neurodegeneration [17, P. 85], [18, P. 319], [19, P. 88]. The involvement of CSTB gene in the general mechanisms that regulate the cell survival [20, P. 561] is also confirmed by the increased lysosomal dysfunction [21, P. 2256], microglial activation [22, P. 40] and axonal degeneration [23, P. e90709] obtained in cystatin B-deficient mouse brains. Cystatin B may be involved in deregulation of autophagy which was found in myoclonus epilepsies [24, P. 991], [8, P. 293], [25, P. 15704]. It is important that PD is also characterized by an activated microglia [26, P. 113] and an impaired autophagy [27, P. 317], [28, P. 728], [29, P. 13].
Together with CSTB proteins, an emerging class of cellular proteins -Arf GAPs (GTPase-activating proteins that induce hydrolysis of GTP bound to ADP-ribosylation factor, also Arf GTPases), is integral component of important signaling pathways [30, P. 347], [31, P. 362 ]. Arf GAPs participate in dendritic differentiation [32, P. 2683], cytoskeleton reorganization [33, P. 792], membrane remodeling and trafficking due to specific interactions with phospholipids and proteins [34, P. 541], [35, 258], [36, P.329] as well as secretion, endocytosis, phagocytosis, cytokinesis, cell adhesion [37, P. 2646], [38, P. 59]. Like CSTB proteins, the Arf GTPases have been shown to play key roles in autophagy [39, P. 1059]. Thus, along with other factors, the disturbances in enzymatic activities of Arf GAPs leading to destruction of membrane remodeling processes may be reasons of human disease [40, P.624], as it was shown in some forms of PD [41, P. S92], [42, P. 1074], [43, P. e1002526].
We previously showed that the gene ACAP3 (ArfGAP encodes centaurin β-5) may be connected with epilepsy and PD. Polymorphism of intronic minisatellite UPS29 of gene ACAP3 have been associated with onset of PD and with some forms of epilepsy in women [44, P. 128], [45, P. 33]. At the same time, this minisatellite exhibited an enhancer-like activity in neuronal type cells [46, P. 715]. It is possible, that disturbances in this minisatellite referred to these pathologies may be reflected in the change of the ACAP3 gene expression.
Taken together, these data indicate a potential common metabolic pathways in some forms of epilepsy and PD and a possible involvement of ACAP3 and CSTB genes in pathogenesis of these diseases. We suppose that these two genes jointly participate in metabolic network of epilepsy and PD disorders and coexisting changes in their expression could be expected. Moreover, we assume that involvement of CSTB gene will be more common sign of other forms of epilepsy along with a clearly defined mutations of this gene in the pathogenesis of EPM1.
Thus, the aim of this study was to detect the mRNA levels of ACAP3 and CSTB genes in leukocytes of periphery venous blood of patients with symptomatic epilepsy and PD.
Methods
Subjects (patients and control group)
Informed written consent was obtained from all participants according to the ethical committee of the Institute of Experimental Medicine RAMS. All the patients (from Neurologic Clinic of the Institute of Experimental Medicine) and controls were residents of St-Petersburg (Russian Federation) and Caucasian. The healthy subjects included 11 men (M ± SD, 34,1 ± 14,1 years old) and 21 women (M ± SD, 30,3 ± 10,1 years old); the PD subjects included 9 men (M ± SD, 41,0 ± 9,2 years old) and 10 women (M ± SD, 42,4 ± 9,9 years old), and the epilepsy subjects included 11 men (M ± SD, 36,5 ± 16,6 years old) and 11 women (M ± SD, 37,1 ± 14,3 years old). Symptomatic epilepsies (polymorphous convulsive and partial attacks) are easily diagnosed by the presence of developmental delay, neurologic abnormalities, and a characteristic seizure and EEG pattern. PD (early onset (30 –50 years of age) diagnoses are based on the combination of the clinical motor signs of bradykinesia, rigidity, tremor and postural instability and non-motor symptoms such as depression and apathy. Healthy controls had no apparent neurological symptoms, had no history of seizures, epilepsy, personal or family neurodegenerative diseases.
Isolation of leukocyte mass
Venous blood (5 ml) was collected into "K+ -EDTA" tubes (Sarstedt, Germany) and was incubated at 37 °C for 1 hour. Plasma with a layer of leukocyte mass was transferred to 2 ml tubes and centrifuged at 100 g for 10 min. The supernatant (serum) was removed; the leukocyte pellet was resuspended in 200 μl of 1×PBS. The leukocyte concentration was evaluated in the Goryaev’s chamber. About 3x106 cells were taken for the subsequent isolation of total RNA from the leukocyte mass, centrifuge at 100 g for 5 min. Supernatant (1XPBS) was removed, the leukocyte pellet was resuspended in 500 μl of TRIzol reagent (Invitrogen, USA). The suspension of leukocytes in TRIzol reagent was stored at -20 0C until RNA isolation, but not more than 1 month.
Isolation of total RNA
Leukocyte mass (3x106 cells) in 500 mkl TRIzol reagent was thawed, another 500 μl of Trisol was added, mixed and incubated at room temperature for 15 minutes. Then total RNA was extracted from these cells according to manufacturer’s instructions (Invitrogen, USA):
The samples were centrifuged at 12.000 g for 10 minutes at 40C. Supernatant (TRIsol fraction) was transferred to a new 1.5 ml centrifuge tube, and 200 μl of cooled chloroform: isoamyl alcohol mix (24:1) was added, carefully mixed and incubated at room temperature for 15 minutes. The samples were centrifuged at 12.000 g for 15 minutes at 40C. The upper phase (containing RNA) was transferred to a new 1.5 ml tube, and 500 μl of cooled isopropanol was added, carefully mixed and incubated for 10-15 minutes at room temperature. The samples were centrifuged at 12.000 g for 15 minutes at 40C. Supernatant was removed. 800 μl of 70% ethanol was added to the RNA precipitate. 70% ethanol was prepared on distilled ethanol and DEPC-treated H2O. Incubate 5 min at room temperature. The samples were centrifuged at 12.000 g for 15 minutes at 40C. The step with 70% ethanol was repeated once more. The samples were centrifuged at 12.000 g for 15 minutes at 4 0C. Supernatant was removed; the RNA precipitate was dried in air from alcohol residues. RNA precipitate was dissolved in 30-50 μl of deionized diethyl-pyrocarbonate-treated water (0,1% DEPC) (Median Filter, Russia). RNA concentration was determined using the Nanodrop 2000c spectrophotometer (Thermo Scientific, USA). Extracted RNA quality was assessed with electrophoresis in a 1,2% agarose gel.
DNase treatment
DNase treatment of RNA solution was carried out for 30 min at 37°C using 2 units of DNase I (Qiagen, Germany), 2 units of RNase inhibitor (Synthol, Russia), total volume of 30 µl. DNase was inactivated by adding EDTA to a final concentration of 2 mM. The DNase-treated RNA was precipitated by the addition of NaCl (final concentration 0.2 M) and 2.5 volumes of distilled ethanol. Samples were incubated for night at -200C, and then were centrifuged for 15 minutes at 12.000 g. Supernatant was removed, 500 μl of 70% ethanol was added to the precipitate, and at 12.000 g for 15 minutes at 40C. Supernatant was removed, and the precipitate was dried in air from alcohol residues. RNA was dissolved in 20 μl of deionized diethyl-pyrocarbonate-treated water (Median Filter, Russia). RNA concentration was determined using the Nanodrop 2000c spectrophotometer (Thermo Scientific, USA). Extracted RNA quality was assessed with electrophoresis in a 1,2% agarose gel. The presence of DNA impurities in the RNA solution was checked by real-time PCR with primers to exons 17 and 18 of ACAP3 gene. In the case of a positive PCR response, the DNase treatment was repeated.
Quantative Reverse Transcription-PCR (QRT-PCR)
DNA-free RNA samples were reverse transcribed using the iScript ™ cDNA Synthesis Kit (Bio-Rad, USA) according to manufacturer's instructions.
Reverse transcription was carried out in 25 μl mix (1x reaction buffer, 0.8 μg total RNA, 1 μl M-MLV reverse transcriptase, oligo (dT) primers): 25ºC - 5 min, 42ºC - 30 min, 85ºC - 5 min. The resulting cDNA was stored at -20 ºC for no more than 1 month. 2 μl of the obtained cDNA was used for real-time PCR. QRT-PCR was performed with the CFX96 Thermal cycler (Bio-Rad, USA) using the iQ ™ SYBR ® Green Supermix Kit (Bio-Rad, USA), by adding 0.88 pmol of each primer (Table 1) and 2 µl of cDNA, the final volume of 25 μl. Each sample was amplified in triplicate at following conditions: denaturation at 95°C for 5 min, followed by 40 cycles consisting of denaturation 95°C for 20 sec, annealing 57°C - 61°C (depending on the target cDNA) for 20 sec, elongation 72°C for 30 sec. PCR specificity was evaluated by amplification product melt curves generated by heating at 0.5°C increments at a rate of 2 sec/step over a temperature range of 74°C - 93°C. Negative controls included DNA-free RNA for each sample, reaction mixture without matrix. Amplification efficiencies for primers were calculated and ranged from 95 % to 105 % mRNA level for ACAP3 and CSTB genes were normalized to the two house-keeping genes ACTB and B2M, one of the most stable expressed genes in white blood cells.
Table 1 – QRT-PCR oligonucleotide primers
| Gene Name | Primer sequence | Tannealing, 0С |
| ACTB | Forward; tccctggagaagagctacg | 61 |
| Reverse: gtagtttcgtggatgccaca | ||
| B2M | Forward: gatgagtatgcctgccgtgt | 57 |
| Reverse: caatccaaatgcggcatct | ||
| ACAP3 | Forward: tgcaggcagcaggaaacc | 57 |
| Reverse: ccacgtatttgtccttgatcca | ||
| CSTB | Forward: atcgccgaccaggtgagg | 59 |
| Reverse: ctcgcaggtgtacgaagtcc |
Statistical analysis of data
The correspondence of the obtained quantitative data to the law of normal distribution was checked using Shapiro-Wilk's criterion (W) for small samples. Non-parametric Kruskal-Wallis test (H) was used to compare the three samples since in all the studied groups the obtained data did not correspond to the normal distribution. The differences were considered statistically significant at p <0,05. Then, pairwise comparisons (for independent groups) were performed using a non-parametric Dunn test (Q) with the Bonferroni correction for the multiplicity of comparisons. In this case the differences were considered statistically significant at p <0,017. Quantitative values are presented in the form of a median and interquartile range (the 25th and 75th percentiles).
Results and Discussion
The problem of identifying new molecular biomarkers of epilepsy and PD with the use of biological material that does not require serious invasive intervention has been widely discussed. Unlike brain biopsy, which allows analyzing the patient condition only in a strictly defined period of the disease development, tests based on the use of peripheral venous blood can enable the most successful selection of individual treatment, monitor the effectiveness of therapy and the risk of resumption of seizures. In addition, the use of such biomarkers in the search for and verification of new antiepileptic and antiparkinsonism drugs will reduce the cost of creating new medicines.
The aim of this study was to quantify the mRNA of the ACAP3 and CSTB genes in the white blood cells (in the total leukocyte mass) at symptomatic epilepsy and early onset PD in order to identify new (potential) biomarkers of these neurological diseases for the development of rapid and informative molecular test-systems suitable for clinical diagnostics.
In this study we revealed the statistically significant changes of mRNA level of ACAP3 and CSTB genes in leukocyte mass from peripheral venous blood of patients with both symptomatic epilepsy and PD using QRT-PCR, while there were gender differences in the level of expression of the studied genes (Fig. 1,2; Table 2).
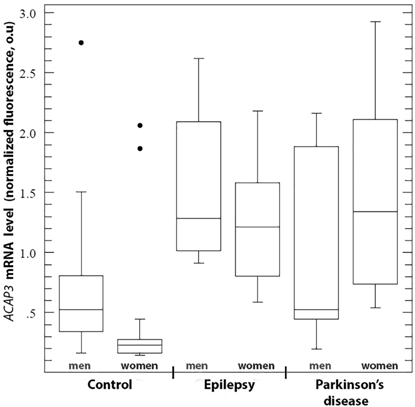
Fig.1 – ACAP3 mRNA level in leukocytes of periphery venous blood of patients with symptomatic epilepsy and early onset Parkinson’s disease
The data are presented as a median and interquartile range (the 25th and 75th percentiles). It is showed the minimum and maximum values. The points indicate the values of emissions from the sample
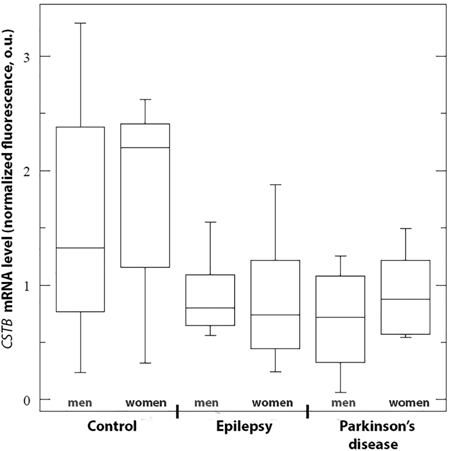
Fig. 2 – CSTB mRNA level in leukocytes of periphery venous blood of patients with symptomatic epilepsy and early onset Parkinson’s disease
The data are presented as a median and interquartile range (the 25th and 75th percentiles). It is showed the minimum and maximum values
Table 2 – The results of statistical analysis of data using non-parametric tests
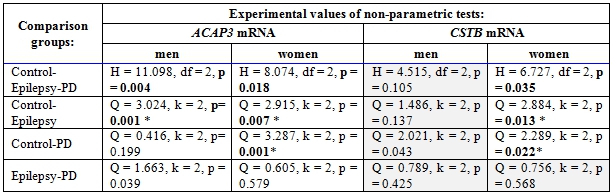
Comments: H - Kruskal-Wallis chi-squared statistic, df- degrees of freedom, Q - Dunn statistica, k - comparison group number, p - p-value; * - statistically significant difference (considering the Bonferroni correction).
In the studied sample of patients with epilepsy, the level ACAP3 mRNA was increased approximately 2.5-fold in men and 5-fold in women compared with the control group. At the same time in sample of PD patients, an increase (about 6-fold) in the level ACAP3 expression was observed only in women. There were no statistically significant differences ACAP3 mRNA level between groups of patients with epilepsy and PD. On the contrary, for CSTB gene statistically significant decrease of mRNA level was observed only in women with epilepsy (about 3-fold) and PD (about 2.5-fold), whereas in men CSTB mRNA level did not differ from control. It should be noted that in both epilepsy and PD there were no gender differences in ACAP3 and CSTB expression within the group, but we observed statistically significant differences in the ACAP3 mRNA level between men and women in the control sample between.
Observed by us a considerable decrease of CSTB mRNA level for symptomatic epilepsy is consistent with data of significant down regulation of this gene expression shown for EPM1 and in CSTB-deficient mice model [15, P. 1731], [47, P. 251], [48, P. 791], [49, P.298], [50, P. 185]. Moreover, we found the same changes of CSTB gene expression for PD. These data also suggest the possible involvement of cystatin B in neurodegenerative processes in Parkinson's disease. Cystatin B as cysteine proteinase inhibitors promotes a survival of neuronal cells and defends them against adverse proteolysises [19, P. 88]. Also, cystatin B is closely involved in abnormal protein metabolism related to amyloid formation [17, P. 85]. It has been suggested that the levels of some cystatins in tissue and body fluids can serve as relatively reliable markers for a variety of diseases [10, P.5406]. The similar downregulation of the CSTB gene expression shown in our study, evidence in favor of the common metabolic pathways leading to neurodegeneration in epilepsy and PD. Neuroplasticity may be another overlapping mechanism of PD and epilepsy [6, P.349]. Epilepsy and PD are often manifested the common symptoms including the cognitive impairment and depression [9, P. 61]. Also, an altered noradrenergic nervous system is known to be involved in neurologic conditions such as epilepsy (mainly, temporal lobe epilepsy) and PD [51, P. 213], [9, P. 61]. Proteomic analysis in a rat hippocampal model showed a link between modified proteins expression (including cystatin B) with the noradrenaline level changed by drugs action [52, P. 1243]. Such post-translational protein modification may be involved in modulation of neuronal excitability during these diseases [53, P. 4740] together with the deregulation of ArfGAPs-coupled exocytotic neurotransmitter release and membrane traffic [54, P. 7919], [55, P. 1143]. Furthermore, brain hyperexcitability in autosomal-recessive idiopathic epilepsy is connected with cytoarchitectural alterations mediated via ArfGAP6 [56, P. 365].
We found the significant upregulation of ACAP3 gene expression in white blood cells of patients with epilepsy and PD, and at the same time gender differences in ACAP3 mRNA level were observed. . The gender differences are seen in some cases of PD, namely, severity of non-motor symptoms are more presented in women with PD [57, P. 33]. Likewise, in women of different populations the single nucleotide polymorphism in some genes is associated with PD [57, P. 33], [58, P. 312]. Previously, we also showed that short alleles of minisatellite UPS29 of ACAP3 gene are associated with PD only for women [44, P. 128], [45, P. 33]. Perhaps, the gender dependence of ACAP3 gene expression in PD revealed by us may also be connected with localization of ACAP3 gene in imprinted chromosomal cluster 1p36 [59, P. 183]. Indeed, similar sex-dependent bias in expression of some imprinted gene in brain was earlier shown [60, P. 682], [61, P. e3839].
It should be noted that there are several works devoted to the search for genes, which had the expression change in white blood cells in PD and epilepsy. Thus, it was observed the overlap in the expression of some genes between the brain and leukocytes in PD [62. P. 19]. Moreover unique patterns of gene expression in white blood cells (total all leukocytes) are described in children with epilepsy, who received treatment with valproic acid and carbamazepine [63, P. 159] as well as in children with idiopathic epilepsy, who not treated, but had differences in expression at partial and generalized seizures [64, P.272].
For example, a decrease of ACAP3 expression was found in peripheral blood leukocytes at symptomatic and presymptomatic patients with Huntington's disease [65, P. 11023] (NCBI GEO database, accession GDS1332).
In addition, GeneCards database (www.genecards.org) shows the same mRNA profile of the ACAP3 (GCID: GC01M001292) and CSTB (GCID: GC21M043772) genes for the brain and leukocytes in normal human tissues (microarray and RNAseq results).
Taking into account that differential genomic expression patterns for each mechanism of injury are common to both brain and blood [58], we may assume that two new candidate of genes expressed in the blood define the abnormalities in protein pathways in brain tissue of patients with symptomatic epilepsy and early-onset PD.
Conclusions
Thus, we demonstrate specific mRNA expression patterns of two genes in leukocyte mass from periphery blood of patients with symptomatic epilepsy and PD which differ from the same of healthy controls. Our data concerned the simultaneous changes of ACAP3 and CSTB gene expression confirm an possible involvement of these genes in the epilepsy and PD neuropathology.
In general, demonstration of the ACAP3 and CSTB gene expression in white cells blood of patients with symptomatic epilepsy and/or PD would be important both for investigation into pathophysiology of these diseases and for the development of potential biomarkers. Of course, in theory it cannot be excluded the secondary nature of the detected changes in mRNA levels of these genes in leukocytes as a result of the effects of pharmacological drugs. But in this case, it should be noted that in the treatment of epilepsy and PD there were used pharmacological preparations with different biochemical (physiological) effects but the observed here effect (change in ACAP3 and CSTB mRNA levels) was almost identical for epilepsy and PD. This fact may suggest that the nature of the therapeutic treatment is not the main factor affecting the expression of ACAP3 and CSTB genes in leukocytes in these diseases.
Список литературы / References
- Ono T. Epilepsy and epileptic syndrome / T. Ono, A. S. Galanopoulou // Adv. Exp. Med. Biol. – 2012 - Vol. 724. – P. 99-113.
- von Bohlen O. Genes, proteins, and neurotoxins involved in Parkinson’s disease / O. von Bohlen, A. Schober, K. Krieglstein // Prog. Neurobiol. – 2004. – Vol. 73. – P. 151–177.
- Nakamura T. Redox regulation of protein misfolding, mitochondrial dysfunction, synaptic damage, and cell death in neurodegenerative diseases / T. Nakamura, D. H. Cho, S. A. Lipton // Exp. Neurol. – 2012. – Vol. 238(1). – P. 12-21.
- Bodenmann P. Neurological comorbidity in Parkinsonism / P. Bodenmann, J. Ghika, G. Van Melle and others // Rev. Neurol. (Paris). – 2001. - Vol.157(1). – P. 45-54.
- Gitatzis A. The epidemiology of the comorbidity of epilepsy in the general population / A. Gitatzis, K. Carrol, A. Majeed and others // Epilepsia. – 2004. – Vol.45(2). – P. 1613- 1622.
- Feddersen B. Parkinson’s disease: Less epileptic seizures more status epilepticus / B. Feddersen, J. Rémi, M. Einhellig and others // Epilepsy Res. – 2014. – Vol. 108(2). – P. 349-354.
- Puschmann A. Alpha-synuclein multiplications with parkinsonism, dementia or progressive myoclonus? / A. Puschmann, Z.K. Wszolek, M. Farrer and others // Parkinsonism Relat. Disord. – 2009. – Vol.15(5). – P. 390-392.
- Polajnar M. Impaired autophagy: a link between neurodegenerative diseases and progressive myoclonus epilepsies / M. Polajnar, E. Zerovnik // Trends Mol. Med. – 2011 – Vol.17(6). – P. 293-300.
- Szot S. Common factors among Alzheimer’s disease, Parkinson’s disease, and epilepsy: Possible role of the noradrenergic nervous system / S. Szot // Epilepsia. – 2012. – Vol. 53. P. 61–66.
- Turk V. Cystatins: biochemical and structural properties, and medical relevance / V. Turk, V. Stoka, D. Turk // Front. Biosci. – 2008. – Vol.13. – P. 5406-5420.
- Di Giaimo R. New insights into the molecular basis of progressive myoclonus epilepsy: a multiprotein complex with cystatin B / R. Di Giaimo, M. Riccio, S. Santi and others // Hum. Mol. Genet. – 2002. – Vol.11. – P. 2941–2950.
- Joensuu T. Molecular background of EPM1-Unverricht-Lundborg disease / T. Joensuu, A.E. Lehesjoki, O.Kopra // Epilepsia. – 2008. – Vol. 49(4). – P. 557-563.
- Ulbrich L. Cystatin B and SOD1: Protein-protein iteraction and possible relation to Neurodegeneration / L. Ulbrich, M. Cozzolino, E. S. Marini and others // Cell Mol. Neurobiol. – 2014. – Vol.34(2). – P.205-213.
- Lalioti M.D. Dodecamer repeat expansion in cystatin B gene in progressive myoclonus epilepsy / M.D. Lalioti,, H.S. Scott, C. Buresi and others // Nature. - 1997. - Vol.386. - P. 847−851.
- Pennacchio L.A. Mutations in the gene encoding cystatin B in progressive myoclonus epilepsy (EPM1) / L.A. Pennacchio, A.E. Lehesjoki, N.E. Stone and others // Science. – 1996. -Vol.271(5256). - P. 1731-1734.
- Lehtinen M.K. Cystatin B deficiency sensitizes neurons to oxidative stress in progressive myoclonus epilepsy, EPM1 / M.K. Lehtinen, S. Tegelberg, H. Schipper and others // J. Neurosci. - 2009. - Vol.29(18). - P. 5910-5915.
- Anderluh G. Pore formation by human stefin B in its native and oligomeric states and the consequent amyloid induced toxicity / G. Anderluh, E. Zerovnik // Front. Mol. Neurosci. – 2012. - Vol.5. - P. 85.
- Ceru S. Intracellular aggregation of human stefin B: confocal and electron microscopy study / S.Ceru, R. Layfield, T. Zavasnik-Bergant and others // Biol. Cell. – 2010. - Vol.102(6). - P. 319-334.
- Polajnar M. Human stefin B normal and patho- physiological role: molecular and cellular aspects of amyloid-type aggregation of certain EPM1 mutants / M. Polajnar, S. Ceru, N. Kopitar-Jerala and others // Front Mol. Neurosci. – 2012. - Vol.5. - P. 88.
- Ganesh S. Neuronal survival in epilepsy: to die or not to die? / S. Ganesh, S. Singh // J. Biosci. – 2005. - Vol.30(5). - P. 561-566.
- Kaur G. Cystatin C rescues degenerating neurons in a cystatin B- knockout mouse model of progressive myoclonus epilepsy / G. Kaur, P. Mohan, M. Pawlik and others // Am. J. Pathol. – 2010. - Vol.177(5). - P. 2256-2267.
- Tegelberg S. Early microglial activation precedes neuronal loss in the brain of the Cstb-/- mouse model of progressive myoclonus epilepsy, EPM1 / S. Tegelberg, O. Kopra, T. Joensuu and others // J. Neuropathol. Exp. Neurol. – 2012. - Vol.71(1). - P. 40-53.
- Manninen O. Progressive volume loss and white matter degeneration in cstb-deficient mice: a diffusion tensor and longitudinal volumetry MRI study / O. Manninen, T. Laitinen, K.K. Lehtimäki and // PLoS One. – 2014. - Vol.9(3). - P. e90709.
- Knecht E. Impaired autophagy in Lafora disease / E. Knecht, C. Aguado, S. Sarkar and others // Autophagy. – 2010. - Vol.6(7). - P. 991-993.
- McMahon J. Impaired autophagy in neurons after disinhibition of mammalian target of rapamycin and its contribution to epileptogene / J. McMahon, X. Huang, J. Yang and others // J. Neurosci. – 2012. - Vol.32(45). - P. 15704-15714.
- Sanchez-Guajardo V. Neuroimmunological processes in Parkinson's disease and their relation to α-synuclein: microglia as the referee between neuronal processes and peripheral immunity / V. Sanchez-Guajardo, C.J. Barnum., M.G. Tansey and others // ASN Neurol. – 2013. - Vol.5(2) - P. 113-139.
- Cheung Z.H. Autophagy deregulation in neurodegenerative diseases - recent advances and future perspectives / Z.H. Cheung, N.Y. Ip // J. Neurochem. – 2011. - Vol.118(3) - P. 317-325.
- Mizushima N. Autophagy: renovation of cells and tissues / N. Mizushima., M. Komatsu // Cell. – 2011. - Vol. 147(4). - P. 728-741.
- Xiong N. The role of autophagy in Parkinson's disease: rotenone-based modeling / N. Xiong, J. Xiong, M. Jia and others // Behav. Brain Funct. – 2013. - Vol.9. - P. 13.
- D'Souza-Schorey C. ARF proteins: roles in membrane traffic and beyond / C. D'Souza-Schorey, P. Chavrier // Nat. Rev. Mol. Cell Biol. – 2006. - Vol.7(5). - P. 347-358.
- Donaldson J.G. ARF family G proteins and their regulators: roles in membrane transport, development and disease / J.G. Donaldson, C.L. Jackson // Nat. Rev. Mol. Cell Biol. – 2011. - Vol.12(6). - P. 362-375.
- Moore C.D. The neuronal Arf GAP centaurin alpha1 modulates dendritic differentiation / C.D. Moore, E.E. Thacker, J. Larimore and others // J. Cell Sci. – 2007. - Vol.120(Pt 15). - P. 2683-2693.
- Venkateswarlu K. PI-3-kinase-dependent membrane recruitment of centaurin-alpha2 is essential for its effect on ARF6-mediated actin cytoskeleton reorganization / K. Venkateswarlu, K.G. Brandom, H.Yun // J. Cell Sci. – 2007. - Vol.120(Pt 5). - P. 792-801.
- Thacker E. The arf6 GAP centaurin alpha-1 is a neuronal actin-binding protein which also functions via GAP-independent activity to regulate the actin cytoskeleton / E. Thacker, B. Kearns, C. Chapman and others // Eur. J. Cell Biol. – 2004. - Vol.83. - P. 541–554.
- Campa F. Arf GTPase-activating proteins and their potential role in cell migration and invasion / F. Campa, P.A. Randazzo // Cell Adh. Migr. – 2008. - Vol.2(4). - P. 258-262.
- Bui Q.T. Large Arf1 guanine nucleotide exchange factors: evolution, domain structure, and roles in membrane trafficking and human disease / Q.T. Bui, M.P. Golinelli-Cohen, C.L. Jackson // Mol. Genet. Genomics. - Vol.82(4). - P. 329-350.
- Spang A. Arf GAPs: gatekeepers of vesicle generation / A. Spang, Y. Shiba, P.A. Randazzo // FEBS Lett. – 2010. - Vol.584(12). - P. 2646-2651.
- Hongu T. Activation machinery of the small GTPase Arf6 / T. Hongu, Y. Kanaho // Adv. Biol. Regul. – 2014. - Vol.54. - P. 59-66.
- Bento C.F. The role of membrane-trafficking small GTPases in the regulation of autophagy / C.F. Bento, C. Puri, K. Moreau and others // J. Cell. Sci. - 2013. - Vol.126(Pt 5) . - P. 1059-1069.
- Dani N. ADP-ribosylated proteins as old and new drug targets for anticancer therapy: the example of ARF6 / N. Dani, A.J. Barbosa, A. Del Rio and others // Curr. Pharm. Des. - 2013. - Vol.19(4) . - P. 624-633.
- Moore D.J. The biology and pathobiology of LRRK2: implications for Parkinson's disease / D.J. Moore // Parkinsonism Relat. Disord. - 2008. - Suppl Vol.2. - P. S92-98.
- Xiong Y. LRRK2 GTPase dysfunction in the pathogenesis of Parkinson's disease / Y. Xiong, V.L. Dawson, T.M.Dawson // Biochem. Soc. Trans. - 2012. - Vol.40(5). - P. 1074-1079.
- Stafa K. GTPase activity and neuronal toxicity of Parkinson's disease-associated LRRK2 is regulated by ArfGAP1 / K. Stafa, A. Trancikova, P.J. Webber and others // PLoS Genet. - 2012. - Vol.8(2). - P. e1002526.
- Suchkova I.O. Analysis of the Association of Minisatellite UPS29 of CENTB5 Gene with Parkinson’s Disease / I.O. Suchkova, D.M. Shubina, A.F. Yakimovskii and others // Russian J. Genetics: Applied Res. - 2011. - Vol. 1 (2). - P. 128–137.
- Pitcik E.V. Association of minisatellite UPS29 with symptomatic and cryptogenic epilepsy in women / E.V. Pitcik, I.O. Suchkova, I.V. Milyukhina and others // Eur. J. Med. Res. - 2011. - Vol.A. – P. 33.
- Sasina L.K. Human intra-intronic minisatellite UPS29 associated with neurological diseases regulates reporter gene EGFP expression depending on cell type / L.K. Sasina, N.A. Slominskaia, I.O. Suchkova and others // Tsitologiia. - 2010. - Vol.52(9) - P. 715-723.
- Pennacchio L.A. Progressive ataxia, myoclonic epilepsy and cerebellar apoptosis in cystatin B-deficient mice / L.A. Pennacchio, D.M. Bouley, K.M. Higgins and others // Nat. Genet. - 1998. – Vol. 20. - P. 251−258.
- Lalioti M.D. Altered spacing of promoter elements due to the dodecamer repeat expansion contributes to reduced expression of the cystatin B gene in EPM1 / M.D. Lalioti, H.S. Scott, S.E. Antonarakis // Hum. Mol. Genet. - 1999. - Vol.81. - P. 791– 798.
- Lafreniere R.G. Unstable insertion in the 5' flanking region of the cystatin B gene is the most common mutation in progressive myoclonus epilepsy type 1, EPM1 / R.G. Lafreniere, D.L. Rochefort, N. Chretien and others // Nat. Genet. - 1997. - Vol.15. - P. 298−302.
- Joensuu T. Cystatin B: mutation detection, alternative splicing and expression in progressive myclonus epilepsy of Unverricht-Lundborg type (EPM1) patients / T. Joensuu, M. Kuronen, K. Alakurtti and others // Eur. J. Hum. Genet. - 2007. - Vol.15(2). - P. 185-193.
- Weinshenker D. The role of catecholamines in seizure susceptibility:new results using genetically engineered mice / D. Weinshenker, P. Szot // Pharmacol. Ther. - 2002. - Vol.94. P. 213–233.
- McHugh P.C. Proteomic analysis of rat hippocampus exposed to the antidepressant paroxetine / P.C. McHugh, G.R. Rogers, D.M. Glubb and others // J. Psychopharmacol. - 2010. - Vol.24(8). - P. 1243-1251.
- Walther D.J. Novel roles for biogenic monoamines: from monoamines in transglutaminase-mediated post-translational protein modification to monoaminylation deregulation diseases / D.J. Walther, S. Stahlberg, J. Vowinckel // FEBS J. - 2011. - Vol.278(24). - P. 4740-4755.
- Meyer M.Z. Regulation of neuroendocrine exocytosis by the ARF6 GTPase-activating protein GIT1 / M.Z. Meyer, N. Déliot, S. Chasserot-Golaz and others // J. Biol. Chem. - 2006. - Vol. 281(12). - P. 7919-7926.
- Shiba Y. ArfGAP1 function in COPI mediated membrane traffic: currently debated models and comparison to other coat-binding ArfGAPs / Y. Shiba, P.A. Randazzo // Histol. Histopathol. - 2012. - Vol. 27(9). - P. 1143-1153.
- Falace A. TBC1D24, an ARF6-interacting protein, is mutated in familial infantile myoclonic epilepsy / A. Falace, F. Filipello, V. La Padula and others //Am. J. Hum. Genet. - 2010. - Vol.87(3). - P. 365-370.
- Solla P. Gender differences in motor and non-motor symptoms among Sardinian patients with Parkinson's disease / P. Solla, A. Cannas, F.C. Ibba and others // J. Neurol. Sci. - 2012. - Vol.323(1-2). - P. 33-39.
- San Luciano M. Gender differences in the IL6 -174G>C and ESR2 1730G>A polymorphisms and the risk of Parkinson's disease / M. San Luciano, L. Ozelius, R.B. Lipton and others // Neurosci. Lett. - 2012. - Vol.506(2). - P. 312-316.
- Li D. Interleukin-10 promoter polymorphisms in Chinese patients with Parkinson's disease / D. Li, Q. He, R. Li and others // Neurosci. Lett. - 2012. - Vol. 513(2). - P. 183-186.
- Gregg C. Sex-Specific Parent-of-Origin Allelic expression in the mouse brain / C. Gregg, J. Zhang, J.E. Butler and others // Science. - 2010. - Vol.329. - P. 682-685.
- Wang X. Transcriptome-wide identification of novel imprinted genes in neonatal mouse brain / X. Wang, Q.Sun, S.D. McGrath and others // PLoS One. - 2008. - Vol.3(12). - P. e3839.
- Karlsson M.K. Found in transcription: Accurate Parkinson’s disease classification in peripheral blood / M.K. Karlsson, P. Sharma, J. Aasly, M. and others // J. Parkinson’s Dis. (Research Report). - 2013. - Vol. 3. - P. - 19-29.
- Tang Y. Valproic acid blood genomic expression patterns in children with epilepsy – a pilot study / Y. Tang, T.A. Glauser, D.L. Gilbert and others // Acta Neurol Scand. - 2004. - Vol. 109. - P.159-168.
- Greiner H.M. mRNA blood expression patterns in new-onset idiopathicpediatric epilepsy / H.M.Greiner, P.S. Horn, K.Holland and others // Epilepsia. - 2013. - Vol. 54 (2). - P. 272-279.
- Borovecki F. Genome-wide expression profiling of human blood reveals biomarkers for Huntington's disease / F.Borovecki, L.Lovrecic, J.Zhou and others // Proc Natl Acad Sci USA. - 2005. - Vol. 102 (31). - P. 11023-11028.