Анализ тканеспецифичности экспрессии гена igf1 у годовиков радужной форели
Анализ тканеспецифичности экспрессии гена igf1 у годовиков радужной форели
Аннотация
В исследовании проанализирована тканеспецифичность экспрессии гена igf1 у годовиков радужной форели, выращиваемых в установке замкнутого водоснабжения. Исследование учитывает сезон выращивания рыб, а также температурный режим и условия комбинированного освещения. Экспрессия оценена в печени, мышцах, сердце и мозге методом от-ПЦР-РВ. Установлено, что уровень мРНК igf1 существенно варьирует между тканями: максимальные значения отмечены в печени, тогда как в остальных тканях уровень экспрессии намного ниже. Корреляционный анализ показал тесную взаимосвязь морфометрических показателей, однако их связь с уровнем экспрессии igf1 была слабой или недостоверной. Полученные данные подтверждают тканевую специфичность экспрессии гена igf1 у годовиков радужной форели и отображают уровни экспрессии igf1 в соответствии с организованными условиями выращивания.
1. Введение
Исследование экспрессионного профиля гена igf1 представляет собой актуальную задачу ввиду его прямого участия в росте и развитии рыб в эмбрио- и онтогенезе , , . Уровень циркулирующего инсулиноподобного фактора роста и гормона роста в крови выступает маркером и медиатором роста, интегрируя сигналы от лиганда соматотропина и метаболического статуса. Экспрессия и действие igf1 у радужной форели зависят от сочетания гормональных (GH) и питательных сигналов, а роль igf1 в качестве ключевого медиатора роста подтверждается как на уровне клетки, так и в организме в целом , .
Ген igf1 и его паралог igf2 опосредуют анаболические эффекты соматотропина на рост рыб, при этом перераспределяют между собой нагрузку в зависимости от периода жизни радужной форели и условий ее выращивания. Так, в эмбриогенезе отмечаются высокие уровни мРНК гена igf2, однако более высокая температура инкубации приводит к увеличению экспрессии igf1, а также других генов роста (gh1, igfr1a/b), особенно на стадии выклева . На уровни мРНК гена igf1 в тканях, где возможна его экспрессия, могут влиять различные факторы. При повышении температуры происходит активация транскрипции igf1, что определяет адаптивные возможности эмбриона к изменяющимся условиям окружающей среды уже на ранних стадиях развития. . Так, при осморегуляции, повышается экспрессия igf1 в жабрах, а естественное и искусственное изменение фотопериода не влияет существенно на средние уровни плазменного IGF-1 . При негативных катаболических условиях, таких как голодание, хронический стресс на фоне плотных посадок при выращивании, у радужной форели наблюдаются физиологические и молекулярные изменения, которые ведут к снижению печеночной продукции инсулиноподобного фактора роста или снижению его действия через приобретённую резистентность к соматотропину и/или смену метаболического приоритета от роста к выживанию , . При содержании рыб преднерестового возраста при круглосуточном освещении и ночном кормлении уровень транскрипции igf1 повышается в печени и мышцах , а у молоди радужной форели таких изменений не происходит, более того увеличенный фотопериод вызывает стресс и иммуносупрессию .
Большинство исследований экспрессии инсулиноподобного фактора роста у радужной форели проведено либо в раннем возрасте, либо у зрелых особей. Поэтому в этом исследовании мы сфокусировали внимание на группе рыб годовалого возраста, определили уровни экспрессии гена igf1 и проанализировали возможное влияние внешних факторов на уровни его мРНК.
2. Методы и принципы исследования
Годовики радужной форели породы рофор содержались в Федеральном селекционно-генетическом центре рыбоводства при температуре выращивания 13°С и комбинированном освещении — естественное освещение с 9:00-16:00, искусственный приглушенный свет с 16:00-9:00. Суточная норма кормления на особь составляла 3.5 г. Для исследования были отобраны здоровые рыбы (n=50) в конце февраля. Анальгезия и седация рыб была произведена в отдельном резервуаре с применением трикаина метансульфоната. Были сняты размерно-весовые показатели: масса тела, длина тела до конца хвостового плавника, длина тела до конца чешуйчатого покрова, длина головы, наибольшая высота тела и толщина тела. От каждой особи были взяты образцы ткани сердца, печени, мышечной ткани и мозг целиком. Полученный биоматериал был зафиксирован РНК-стабилизатором IntactRNA («Евроген», Россия). Экстракция тотальной РНК проводилась коммерческим набором «ЛИРА+» («Биолабмикс», Россия). Концентрация и чистота растворенной РНК измерялась спектрофотометром NanoDrop2000с (Thermo Fisher Scientific, США). Синтез кДНК выполнялся с использованием коммерческого набора «M-MuLV-RH» («Биолабмикс», Россия), согласно протоколу производителя. Подбор праймеров выполнялся через онлайн-сервис PrimerBLAST базы данных NCBI (таблица 1).
Таблица 1 - Последовательности праймеров, для секвенирования и оценки относительного уровня экспрессии
Название гена | Последовательность | Длина фрагмента п.н. |
IGF1 | F – GCTTCCAGAGTTGCGAGCTA R – CATGTCTGTGTGGCGTTGTG | 100 |
G6PD | F – TCAAAGAGGGTCGGTTTCCG R – ACCCATCCCTCTTCCTCTCC | 87 |
Для постановки ПЦР в реальном времени использовали готовый микс от компании Биолабмикс «БиоМастер HS-qPCR SYBR Blue(2×)» («БИОЛАБМИКС», Россия) согласно инструкции производителя. Каждый образец ставился в трех повторностях. ПЦР в режиме реального времени проводили на приборе Bio-Rad CFX Opus 96 («Bio-Rad», США, программное обеспечение CFX Maestro Software) по протоколу амплификации: предварительная денатурация при 95,0 °C — 5 минут, далее следовали 40 циклов: 95°C 15 секунд, 60,0 °C 15 секунд, 72,0 °C 30 секунд. Расчет ∆Ct, ∆∆Ct проводились по методике Livak . Для оценки достоверности использовался критерий Краскела-Уоллиса.
3. Основные результаты
Для сопоставления данных об уровнях мРНК гена igf1 с метрическими показателями рыб был выполнен анализ корреляционных связей между размерно-весовыми показателями (Рисунок 1).
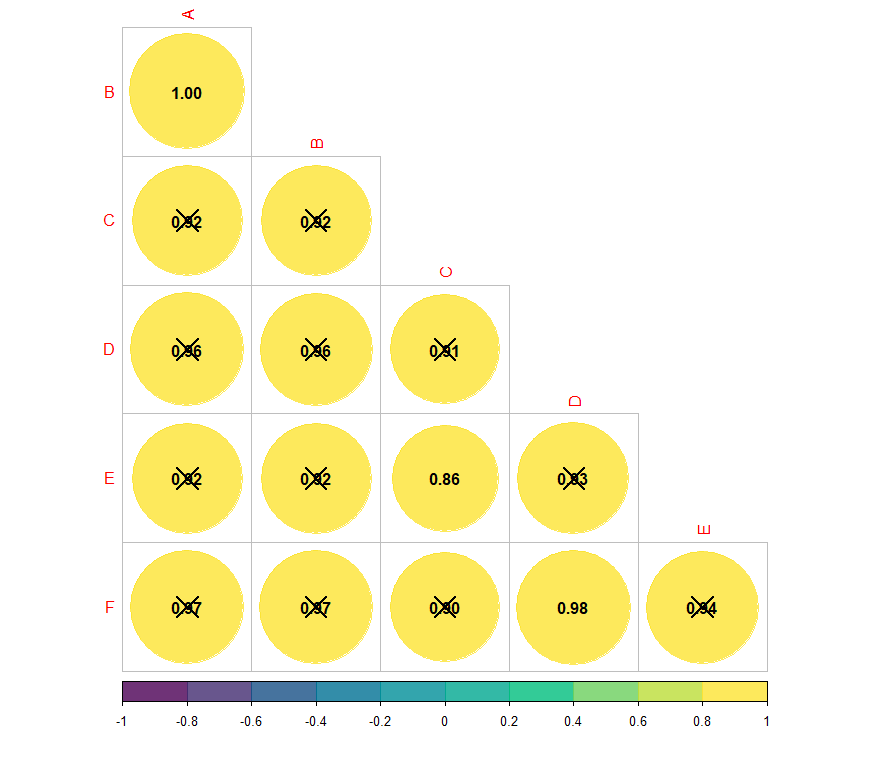
Рисунок 1 - Корреляционные связи размерно-весовых показателей годовиков радужной форели:
А – длина тела по Смиту; B – длина тела до конца чешуйчатого покрова; C – длина головы; D – высота тела; E – толщина тела; F – масса тела; Х – недостоверное значение корреляции
Примечание: цветовая шкала отображает значения корреляции
Всего было собранно метрических данных и биологического материала от 50 особей радужной форели. Результаты анализа относительного уровня экспрессии гена igf1 в мышцах, сердце, печени и мозге представлены на рисунке 2.
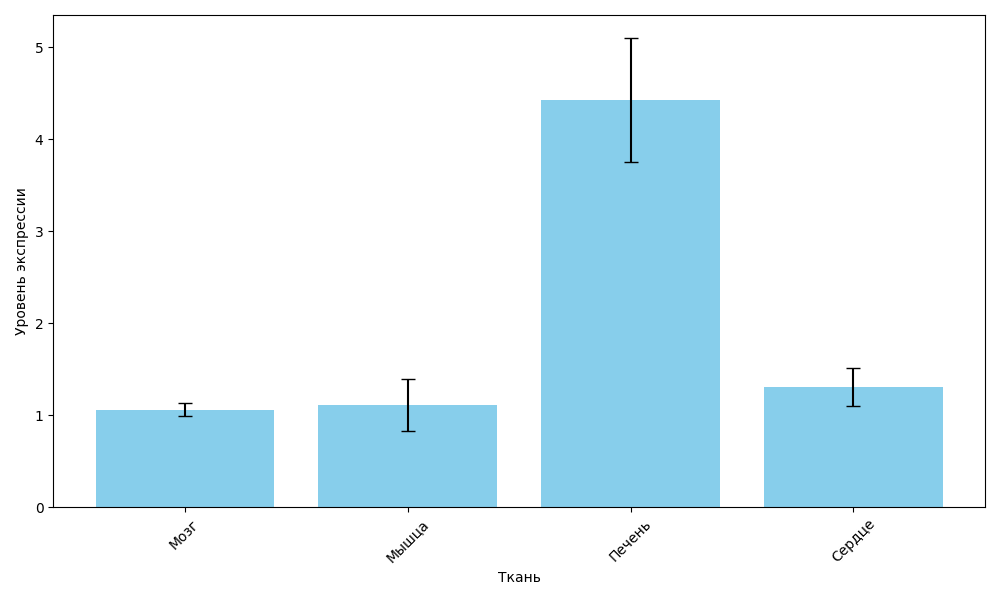
Рисунок 2 - Относительный уровень экспрессии гена igf1 у годовиков радужной форели
Примечание: n=50
Чтобы установить связь уровня экспрессии в разных тканях с метрическими данными был проведен корреляционный анализ (Рисунок 3).

Рисунок 3 - Корреляционные связи размерно-весовых показателей относительного уровня экспрессии у годовиков радужной форели:
А – длина тела по Смиту; B – длина тела до конца чешуйчатого покрова; C – длина головы; D – высота тела; E – толщина тела; F – масса тела; G – относительный уровень экспрессии IGF1 в ткани печени; H – относительный уровень экспрессии igf1 ткани сердца; I – относительный уровень экспрессия igf1 в мышечной ткани; J – относительный уровень экспрессия igf1 в мозге
4. Заключение
Проведенное исследование показало, что у годовиков радужной форели, содержавшихся в зимний период при температуре 13 °C и укороченном естественном фотопериоде в комбинировании с приглушенным искусственным освещением, экспрессия гена igf1 демонстрирует чётко выраженную тканеспецифичность. Максимальный уровень транскрипции был зарегистрирован в печени — основном органе синтеза инсулиноподобного фактора роста у рыб, тогда как в мышцах, сердце и мозге экспрессия оставалась низкой и статистически сравнимой между собой.
Несмотря на высокую взаимосвязанность морфометрических характеристик рыб, их связь с уровнем экспрессии igf1 в разных тканях оказалась неоднозначной. В печени обнаружена средняя отрицательная корреляция, что может отражать снижение чувствительности соматотропной оси или перераспределение метаболических ресурсов при данных параметрах освещения. Положительные корреляции, выявленные в мышцах и мозге, статистически не подтверждены и требуют дальнейшего изучения.
Полученные результаты указывают на то, что в условиях зимнего выращивания (умеренная температура и ограниченный фотопериод) регуляция igf-зависимых процессов у годовиков радужной форели определяется сочетанием сезонных физиологических адаптаций и тканеспецифичных механизмов экспрессии. Эти данные подчеркивают необходимость учитывать температурно-световые условия при интерпретации активности компонентов GH–IGF оси и планировании дальнейших исследований роста рыб в УЗВ.