НОРМАЛИЗАЦИЯ РОСТОВЫХ ХАРАКТЕРИСТИК КЛЕТОК ФИБРОСАРКОМЫ В КУЛЬТУРЕ ПРИ ДЕЙСТВИИ МИТОХОНДРИАЛЬНО-НАПРАВЛЕННЫХ АНТИОКСИДАНТОВ

Титова Е.В.1, Иванова О.Ю.2, Дугина В.Б.3
1ORCID: 0000-0003-4790-6048, 2Кандидат биологических наук, 3Доктор биологических наук, НИИ ФХБ имени А.Н. Белозерского МГУ имени М.В. Ломоносова
НОРМАЛИЗАЦИЯ РОСТОВЫХ ХАРАКТЕРИСТИК КЛЕТОК ФИБРОСАРКОМЫ В КУЛЬТУРЕ ПРИ ДЕЙСТВИИ МИТОХОНДРИАЛЬНО-НАПРАВЛЕННЫХ АНТИОКСИДАНТОВ
Аннотация
Неопластическая трансформация затрагивает изменения морфологии и пролиферации клеток. Известно, что при этом в трансформированных клетках значительно увеличивается уровень внутриклеточных активных форм кислорода (АФК). В ходе настоящего исследования с помощью антиоксидантов (АО) семейства SkQ, адресованных в митохондрии (мАО), была показана роль АФК в изменении ростовых характеристик клеток фибросаркомы НТ1080, в которых ранее был обнаружен активированный онкоген N-ras. Действие мАО на исследуемую клеточную культуру приводило к снижению скорости роста культуры и появлению популяции двуядерных клеток. Наблюдаемое торможение пролиферации сопровождалось появлением митотических клеток с аномальными морфологическими характеристиками, задержкой на стадии цитокинеза. Таким образом, настоящее исследование поднимает вопрос о возможном использовании мАО для ингибирования пролиферации клеток с Ras-трансформированным фенотипом.
Ключевые слова: неопластическая трансформация, активированный N-ras онкоген, пролиферация клеток, антиоксиданты.Titova E.V.1, Ivanova O.Ju.2, Dugina V.B.3
1ORCID: 0000-0003-4790-6048, 2PhD in Biology, 3PhD in Biology, Dr.of Sci, A.N. Belozersky Research Institute of Physico-Chemical Biology MSU
INHIBITION OF PROLIFERATION OF FIBROSARCOMA CELLS IN CULTURE BY MITOCHONDRIA-TARGETED ANTIOXIDANTS
Abstract
Neoplastic transformation is leading to morphological and proliferative cell disorders. Increased level of intracellular reactive oxygen species (ROS) in transformed cells is well established. Here we have shown the role of ROS in stimulation of cell proliferation of HT1080 fibrosarcoma cells with activated N-ras oncogene. We have found that mitochondria-targeted antioxidant SkQ1 and its analogs inhibit cell growth and increase binuclear cell population. On the other hand, the treatment with SkQs induced pathological changes in mitotic cells morphology and cytokinesis lead-time. We concluded that our finding put the question whether the mitochondria-targeted antioxidants can be used for inhibiting proliferation of tumor cells with Ras-induced morphological phenotype.
Keywords: neoplastic transformation, activated N-ras oncogene, cell proliferation, antioxidants.Введение.
Активные формы кислорода (далее АФК) непрерывно генерируются в клетках и являются важными посредниками во многих сигнальных каскадах, участвуя в регуляции таких процессов, как пролиферация и миграция клеток. Многочисленные экспериментальные данные подтверждают роль АФК в процессе неопластической трансформации и опухолевой прогрессии [1], показана усиленная продукция АФК некоторыми опухолевыми клетками [2].
Известно, что основным источником внутриклеточных АФК является митохондрия, где предшественники АФК образуются в электрон-транспортной дыхательной цепи и цикле трикарбоновых кислот [3]. В ряде работ показана тесная связь канцерогенеза с митохондриальными повреждениями [4], поэтому защиту митохондрий клетки от окислительных повреждений можно рассматривать как важную экспериментальную задачу.
На данный момент все большее внимание уделяется изучению эффекта синтетических мАО на патологические состояния клетки.
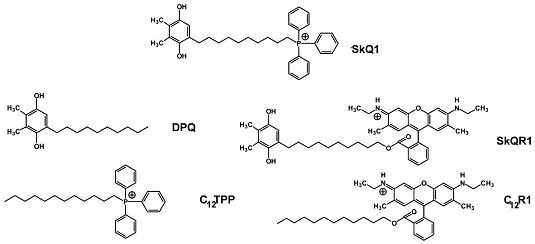
Рис. 1 - Структурные формулы веществ семейства SkQ и его аналогов [5]
В настоящей работе изучался эффект мАО нового класса - SkQ1 и его аналогов (Рис.1). SkQ1 был синтезирован по принципу соединения антиоксидантной молекулы, пластохинона, с проникающим катионом, отвечающим за накопление вещества в митохондриях [6].
Такое патологическое состояние, как неопластическая трансформация, характеризуется увеличением скорости пролиферации, отвечающей за опухолевый рост. Изучение влияния мАО семейства SkQ на пролиферацию и параметры клеточного цикла опухолевых клеток мезенхимального происхождения (фибросаркомы) являлось целью данного исследования.
Методы исследования.
Клетки: линия HT1080 - псевдодиплоидная культура фибросаркомы человека, содержащая мутантный аллель онкогена N-ras [7].
Антиоксиданты: SkQ1 (пластохинонил децилтрифенилфосфониум) и SkQR1 (пластохинонил децилродамин 19, флуоресцентное производное SkQ1) в концентрации 5-100 нМ в среде для культивирования. В качестве контрольных молекул использовали DPQ (децилпластохинон, аналог SkQ1, не содержащий катионную часть), C12TPP (додецилтрифенилфосфониум, аналог SkQ1, не имеющий хиноновую группу) и C12R1 (децилродамин) в тех же концентрациях.
Иммунофлуоресцентная микроскопия: для подсчета клеток на стадии митоза и анализа митотических фаз использовали препараты, меченные DAPI и родамин-меченным фаллоидином, анализировали 500 клеток для каждого условия эксперимента в каждом из трех независимых экспериментов. Подсчет количества клеток при изучении скорости роста производился на единицу площади культуральной чашки (1 мм2) после 48 часов инкубации.
Измерение параметров клеточного цикла методом проточной цитофлуориметрии: распределение клеток по фазам клеточного цикла анализировали по флуоресценции йодида пропидия. Флуоресценцию анализировали с помощью проточного цитофлуориметра Beckman Coulter Cytomics FC500. В каждом образце анализировали не менее 20000 клеток.
Прижизненная фазово-контрастная микроскопия: клетки сажали на покровные стекла в стеклянную камеру. Съемку проводили через 24 часа с помощью цейтраферной видеосъемки (замедление в 120 раз). При подсчете продолжительности фаз митоза было проанализировано 30 митотических клеток для каждого условия эксперимента.
Статистический анализ. Данные представлены как среднее значение ± стандартная ошибка среднего по результатам как минимум трех независимых экспериментов. Для статистической оценки результаты были проанализированы с помощью t-теста Стьюдента. Значения p < 0,001 (***), p < 0,01 (**) и p < 0,05 (*) считались статистически достоверными.
Результаты.
SkQ1 индуцирует снижение пролиферативной активности клеток НТ1080. В ходе данного исследования было показано существенное изменение скорости роста клеток фибросаркомы НТ1080 в присутствии мАО. Так, инкубация с SkQR1 приводила к уменьшению общего количества клеток в культуре более чем в два раза. (Рис. 2, а), гибели клеток при этом не наблюдали. При этом SkQ1 и SkQR1 индуцировали увеличение количества популяции двуядерных клеток в культуре более чем в 10 раз (Рис. 2, б). Контрольные молекулы DPQ, C12ТРР и SkQRC12 не вызывали аналогичных изменений.
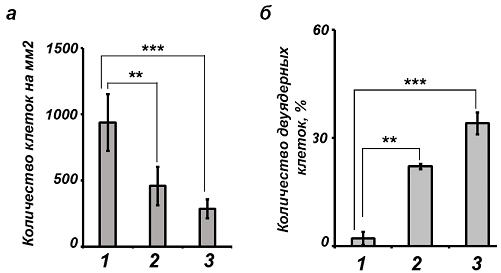
Рис. 2 - Действие мАО (48 часов инкубации) на пролиферацию и плоидность клеток HT1080: а – уменьшение количества клеток в культуре HT1080 после инкубации с SkQR1: 1 - контроль, 2 - SkQR1 (10 нМ), 3 - SkQR1 (20 нМ); б – увеличение количества двуядерных клеток в культуре HT1080 после инкубации с SkQR1 (1 - контроль, 2 - SkQR1 10 нМ, 3 - SkQR1 20 нМ)
Анализ изменений параметров клеточного цикла клеток HT1080 в присутствии мАО методом проточной цитофлуориметрии. При измерении параметров клеточного цикла клеток культуры HT1080 после инкубации с мАО было выявлено уменьшение G1 (2n) популяции и значительное увеличение 4n-популяции (Рис. 3). Увеличение популяции клеток 4n на гистограмме данных проточной цитофлуориметрии могло свидетельствовать как об увеличении относительного количества митотических клеток на стадии G2/M, так и о появлении популяции полиплоидных и многоядерных клеток.
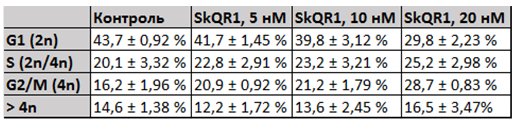
Рис. 3 - мАО индуцируют уменьшение G1 и увеличение G2/M популяции в культуре НТ1080, данные проточной цитофлуориметрии. Изменение параметров клеточного цикла в культуре HT1080, SkQR1 (24 часа инкубации), среднее количество клеток ± стандартная ошибка среднего (%). Статистически значимые различия получены для периодов G1 и G2/M при концентрациях SkQR1 10 нМ (р<0,05) и 20 нМ (р<0,01)
SkQ1 и SkQR1 снижают количество митозов в культуре клеток НТ1080 фибросаркомы человека. Данные проточной цитофлуориметрии были дополнены иммунофлуоресцентным микроскопическим анализом. При исследовании количества митотических клеток в контрольных и обработанных мАО культурах HT1080 было выявлено, что инкубация с SkQR1 и SkQ1 приводит к снижению числа митотических клеток (Рис. 4, а). При этом контрольные молекулы DPQ, C12ТРР и SkQRC12 не оказывали действия на количество митозов.
SkQ1 и SkQR1 индуцируют появление аномальных митотических клеток в культуре HT1080. Для более подробного анализа состава G2/M популяции, выявленной методом проточной цитофлуориметрии, были проведены динамические наблюдения за культурой клеток HT1080. Была выявлена популяция митотических клеток с измененной морфологией на стадии телофазы. Клетки имели неровные контуры и множество активных псевдоподий с многочисленными блеббами. Стадия телофазы в таких клетках (аномальная телофаза) была затянута во времени. Митозы с аномальной телофазой называли аномальными митозами.
В ходе экспериментов было выявлено, что инкубация с SkQ1 и SkQR1 вызывает увеличение продолжительности фаз как нормальных, так и аномальных митозов. Так, длительность аномальной телофазы была в 4,5 - 5 раз дольше, чем в нормальных митотических делениях. После инкубации клеток с мАО продолжительность телофазы как в популяции клеток с нормальными, так и с аномальными митозами увеличивалась в 2 - 2,5 раза. Было также отмечено увеличение продолжительности профазы и метафазы как в нормальных, так и в аномальных митотических делениях.
Помимо этого, в ходе динамических наблюдений было показано, что мАО приводят к значительному увеличению процента аномальных митозов в культуре (Рис. 4, б).
Контрольные молекулы DPQ, C12ТРР и SkQRC12 не влияли на продолжительность фаз митотических делений, увеличения количества аномальных клеточных делений по сравнению с контролем также не наблюдалось.
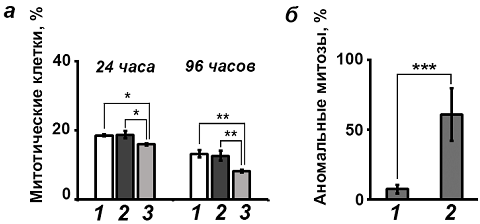
Рис. 4 - Изменение количества митотических делений в культуре клеток НТ1080 под действием мАО: а - действие SkQR1 на количество митотических клеток в культуре НТ1080 (1 - контроль, 2 - SkQRC12, 20 нМ, 3 - SkQR1, 20 нМ, 24 и 96 часов инкубации); б - индукция аномальных митозов в популяции делящихся клеток культуры фибросаркомы НТ1080 после инкубации с мАО (1 – контроль, 2 - SkQ1, 20 нМ, 72 часа)
Таким образом, в ходе исследования было показано значительное увеличение продолжительности митоза клеток НТ1080. Были выявлены: 1) увеличение продолжительности профазы и метафазы после инкубации с SkQ1 и SkQR1; 2) задержка митотических клеток на стадии телофазы. По-видимому, данный эффект мАО на митотические клетки НТ1080 является причиной снижения пролиферативной активности неопластически трансформированных клеток после воздействия на них веществ семейства SkQ.
Обсуждение.
В ходе исследования был показан эффект мАО на трансформированные фибробласты линии НТ1080 с измененным RAS-трансформированным фенотипом. В литературе описано, что экспрессия активированных Ras белков вызывает повышение уровня внутриклеточных АФК за счет переноса электронов на молекулярный кислород НАДФН оксидазой плазматической мембраны и последующей продукции АФК в клетке [8, 9]. В ряде исследований было показано, что SkQ1 и его аналоги эффективно снижают количество внутриклеточных АФК [5, 10]. Таким образом, наблюдаемые эффекты мАО семейства SkQ в большей степени обусловлены снижением количества АФК в клетках и активацией соответствующих сигнальных каскадов.
В ходе настоящей работы было выявлено, что мАО эффективно снижают скорость роста клеток НТ1080, содержащих активированный N-ras онкоген. В ряде исследований показано, что основной ответ клетки на активацию Ras белков - запуск цитоплазматических MAP(Mitogen Activated Protein)-киназных каскадов. Следствием изменений активности транскрипционных факторов, индуцируемых активацией МАР-киназ, является повышение экспрессии гена циклина D1 и увеличение активности циклинзависимых киназ, что стимулирует увеличение скорости пролиферации трансформированных клеток [11]. Описанная в данном исследовании высокая скорость роста клеток культуры НТ1080, которую удалось снизить с помощью мАО семейства SkQ, предположительно может быть обусловлена именно этим сигнальным каскадом.
В ходе настоящей работы с помощью метода проточной цитофлуориметрии было обнаружено увеличение G2/M 4n-популяции в культуре НТ1080 после инкубации с SkQ1 и SkQR1. При этом анализ иммунофлуоресцентно окрашенных препаратов и динамические наблюдения за клетками фибросаркомы выявили снижение пролиферации, задержку клеток на стадии телофазы клеточного деления и увеличение популяции двуядерных клеток в присутствии мАО. Сопоставление полученных данных об ингибировании митотической активности клеток с данными об увеличении G2/M популяции позволило сделать предположение, что 4n-популяцию составляют преимущественно не прошедшие цитокинез полиплоидные и двуядерные клетки.
Суммируя полученные результаты, в ходе исследования была впервые показана роль АФК в изменении пролиферативных характеристик клеток фибросаркомы НТ1080. Экспериментальная модель с участием мАО показала, что АО семейства SkQ эффективно снижают скорость пролиферации клеток. Таким образом, настоящее исследование поднимает вопрос о возможном использовании мАО для ингибирования пролиферации и реверсии нормальных ростовых характеристик опухолевых клеток с Ras-трансформированным фенотипом.
Литература
- Ames B.N. Measuring oxidative damage in humans: relation to cancer and ageing.//IARC Sci Publ.- 1988. - № 89. - c.407-16.
- Szatrowski T.P., Nathan C.F. Production of large amounts of hydrogen peroxide by human tumor cells.//Cancer Res. - - T. 51, № 3. c.794-8.
- Starkov A.A., Fiskum G., Chinopoulos C., Lorenzo B.J., Browne S.E., Patel M.S., Beal M.F.//J Neurosci. - 2004. - T. 24, № 36. - c.7779-88.
- Hail N Jr. Mitochondria: a novel target for the chemoprevention of cancer.//Apoptosis. - 2005. - T. 10, №4. - c.687-705.
- Skulachev V.P., Antonenko Y.N., Cherepanov D.A., Chernyak B.V., Izyumov D.S., Khailova L.S., Klishin S.S., Korshunova G.A., Lyamzaev K.G., Pletjushkina O.Y., Roginsky V.A., Rokitskaya T.I., Severin F.F., Severina I.I., Simonyan R.A., Skulachev M.V., Sumbatyan N.V., Sukhanova E.I., Tashlitsky V.N., Trendeleva T.A., Vyssokikh M.Y., Zvyagilskaya R.A. Prevention of cardiolipin oxidation and fatty acid cycling as two antioxidant mechanisms of cationic derivatives of plastoquinone (SkQs).//Biochim Biophys Acta.- 2010. - T. 1797, № 6-7. - c.878-89.
- AntonenkoN., Avetisyan A.V., Bakeeva L.E., Chernyak B.V., Chertkov V.A., Domnina L.V., Ivanova O.Y., Izyumov D.S., Khailova L.S., Klishin S.S., Korshunova G.A., Lyamzaev K.G., Muntyan M.S., Nepryakhina O.K., Pashkovskaya A.A., Pletjushkina O.Y., Pustovidko A.V., Roginsky V.A., Rokitskaya T.I., Ruuge E.K., Saprunova V.B., Severina I.I., Simonyan R.A., Skulachev I.V., Skulachev M.V., Sumbatyan N.V., Sviryaeva I.V., Tashlitsky V.N., Vassiliev J.M., Vyssokikh M.Y., Yaguzhinsky L.S., Zamyatnin A.A. Jr, Skulachev V.P. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 1. Cationic plastoquinone derivatives: synthesis and in vitro studies.//Biochemistry (Mosc). - 2008. - T. 73, № 12. - c.1273-87.
- Gupta S.,Stanbridge E.J. Pairedhumanfibrosarcomacell lines that possess or lack endogenous mutant N-ras alleles as experimental model for Ras signaling pathways.//Methods Enzymol. - - № 333. - c.290-306.
- Archer H., Bar-Sagi D. Ras and Rac as activators of reactive oxygen species (ROS).//Methods Mol. Biol. - 2002. - № 189. - c. 67–73.
- Seru R., Mondola P., Damiano S., Svegliati S., Agnese S., Avvedimento E.V., Santillo M., HaRas activates the NADPH oxidase complex in human neuroblastoma cells via extracellular signal-regulated kinase 1/2 pathway.// Neurochem. - 2004. - № 91. - c.613–622.
- AntonenkoN., Roginsky V.A., Pashkovskaya A.A., Rokitskaya T.I., Kotova E.A., Zaspa A.A., Chernyak B.V., Skulachev V.P. Protective effects of mitochondria-targeted antioxidant SkQ in aqueous and lipid membrane environments.// J Membr Biol. - 2008. - T. 222, № 3. - c.141–9.
- Campbell S.L.,Khosravi-Far R., Rossman K.L., Clark G.J., Der C.J. Increasing complexity of Ras signaling.//Oncogene. - 2004. - T. 17, № 11. - c.1395-413.
References
- Ames B.N. Measuring oxidative damage in humans: relation to cancer and ageing.//IARC Sci Publ. - 1988. - № 89. - P.407-16.
- Szatrowski T.P., Nathan C.F. Production of large amounts of hydrogen peroxide by human tumor cells.//Cancer Res. - - Vol. 51, № 3. P.794-8.
- Starkov A.A., Fiskum G., Chinopoulos C., Lorenzo B.J., Browne S.E., Patel M.S., Beal M.F.//J Neurosci. - 2004. - Vol. 24, № 36. - P.7779-88.
- Hail N Jr. Mitochondria: a novel target for the chemoprevention of cancer.//Apoptosis. - 2005. - Vol. 10, №4. - P.687-705.
- Skulachev V.P., Antonenko Y.N., Cherepanov D.A., Chernyak B.V., Izyumov D.S., Khailova L.S., Klishin S.S., Korshunova G.A., Lyamzaev K.G., Pletjushkina O.Y., Roginsky V.A., Rokitskaya T.I., Severin F.F., Severina I.I., Simonyan R.A., Skulachev M.V., Sumbatyan N.V., Sukhanova E.I., Tashlitsky V.N., Trendeleva T.A., Vyssokikh M.Y., Zvyagilskaya R.A. Prevention of cardiolipin oxidation and fatty acid cycling as two antioxidant mechanisms of cationic derivatives of plastoquinone (SkQs).//Biochim Biophys Acta.- 2010. - Vol. 1797, № 6-7. - P.878-89.
- AntonenkoN., Avetisyan A.V., Bakeeva L.E., Chernyak B.V., Chertkov V.A., Domnina L.V., Ivanova O.Y., Izyumov D.S., Khailova L.S., Klishin S.S., Korshunova G.A., Lyamzaev K.G., Muntyan M.S., Nepryakhina O.K., Pashkovskaya A.A., Pletjushkina O.Y., Pustovidko A.V., Roginsky V.A., Rokitskaya T.I., Ruuge E.K., Saprunova V.B., Severina I.I., Simonyan R.A., Skulachev I.V., Skulachev M.V., Sumbatyan N.V., Sviryaeva I.V., Tashlitsky V.N., Vassiliev J.M., Vyssokikh M.Y., Yaguzhinsky L.S., Zamyatnin A.A. Jr, Skulachev V.P. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 1. Cationic plastoquinone derivatives: synthesis and in vitro studies.//Biochemistry (Mosc). - 2008. - Vol. 73, № 12. - P.1273-87.
- Gupta S.,Stanbridge E.J. Pairedhumanfibrosarcomacell lines that possess or lack endogenous mutant N-ras alleles as experimental model for Ras signaling pathways.//Methods Enzymol. - - № 333. - P.290-306.
- Archer H., Bar-Sagi D. Ras and Rac as activators of reactive oxygen species (ROS).//Methods Mol. Biol. - 2002. - № 189. - P. 67–73.
- Seru R., Mondola P., Damiano S., Svegliati S., Agnese S., Avvedimento E.V., Santillo M., HaRas activates the NADPH oxidase complex in human neuroblastoma cells via extracellular signal-regulated kinase 1/2 pathway.// Neurochem. - 2004. - № 91. - P.613–622.
- AntonenkoN., Roginsky V.A., Pashkovskaya A.A., Rokitskaya T.I., Kotova E.A., Zaspa A.A., Chernyak B.V., Skulachev V.P. Protective effects of mitochondria-targeted antioxidant SkQ in aqueous and lipid membrane environments.// J Membr Biol. - 2008. - Vol. 222, № 3. - P.141–9.
- Campbell S.L.,Khosravi-Far R., Rossman K.L., Clark G.J., Der C.J. Increasing complexity of Ras signaling.//Oncogene. - 2004. - Vol. 17, № 11. - P.1395-413.