НЕКОТОРЫЕ ЗАКОНОМЕРНОСТИ МОРФОГЕНЕЗА НЕЙРОЦИТОВ ДОМАШНИХ МЛЕКОПИТАЮЩИХ И ПТИЦ
Хохлова С.Н.1, Симанова Н.Г.2, Богданова М.А.3, Степочкин А.А.4, Фасахутдинова А.Н.5
1ORCID:0000-0001-6528-5762, кандидат биологических наук, доцент, 2ORCID: 0000-0003-4713-3380, кандидат биологических наук, доцент, 3ORCID: 0000-0002-1775-5771, кандидат биологических наук, доцент, 4ORCID: 0000-0003-4586-4681, кандидат ветеринарных наук, доцент, 5ORCID: 0000-0001-6743-2367, кандидат биологических наук, доцент, Ульяновская государственная сельскохозяйственная академия
НЕКОТОРЫЕ ЗАКОНОМЕРНОСТИ МОРФОГЕНЕЗА НЕЙРОЦИТОВ ДОМАШНИХ МЛЕКОПИТАЮЩИХ И ПТИЦ
Аннотация
К настоящему времени накопилось много исследований, посвященных изучению постнатального морфогенеза нейроцитов позвоночных. Большинство исследователей единодушно в том, что в постнатальном морфогенезе млекопитающих и птиц, особенно раннем, продолжается формообразование нейроцитов. Также общепризнанным является и вывод о гетерохронии постнатального морфогенеза нейроцитов. Вместе с тем в доступной литературе мы не нашли попыток обобщения накопленных фактов о причинах гетерохронии морфогенеза нейроцитов, его возрастных границах, видовых и индивидуальных особенностях у домашних млекопитающих и птиц. Такие обобщения представляют несомненный интерес для теоретической и клинической нейрологии, а также специалистов - животноводов.
Ключевые слова: узел, морфология, нервные клетки.Khokhlova S.N.1, Simanova N.G.2, Bogdanova M.A.3, Stepochkin A.A.4, Fasakhutdinova A.N.5
1ORCID: 0000-0001-6528-5762 PhD in Biological Sciences, Associate professor, 2ORCID: 0000-0003-4713-3380, PhD in Biological Sciences, Associate professor, 3ORCID: 0000-0002-1775-5771, PhD in Biological Sciences, Associate professor, 4ORCID: 0000-0003-4586-4681, PhD in Veterinary Medicine and Science, Associate professor, 5ORCID: 0000-0001-6743-2367, PhD in Biological Sciences, Associate professor, Ulyanovsk State Agricultural Academy
SOME REGULARITIES OF NERVE CELL MORPHOGENESIS OF DOMESTIC MAMMALS AND BIRDS
Abstract
A lot of works devoted to the study of postnatal nerve cells morphogenesis were conducted for the recent years. The majority of the scientists agree that nerve cells continue the forming in the postnatal morphogenesis of mammals and birds, especially at early stage. The conclusion about the heterochronicity of the postnatal morphogenesis of neurocytes is also generally accepted. At the same time, in the works studied, we have not found any attempts to generalize the accumulated facts about the causes of heterochrony of nerve cells morphogenesis, its age boundaries, species and individual features of domestic mammals and birds. Such generalizations are of undoubted interest for theoretical and clinical neurology, as well as for specialists in cattle breeding.
Keywords: knot, morphology, nerve cells.Настоящее сообщение является попыткой обобщения накопленных более чем за 20 лет в нашей лаборатории данных с учетом имеющихся в литературе.
Нейрогистологическими методами: Бильшовский - Грос, Кампос, Белецкий и др. с использованием элементов морфометрии исследовали постнатальный морфогенез нейроцитов центрального (мотонейроны спинного мозга, клетки Пуркинье мозжечка) и периферического (нейроны спинальных, экстра - и интрамуральных вегетативных ганглиев) отделов нервной системы домашних млекопитающих (крупный рогатый скот, свинья, собака, кролик) и птиц (курица). В качестве оценочных показателей морфогенеза нейроцитов учитывали объем их ядра и цитоплазмы, величину ядерно-плазменного отношения, морфологию дендритного аппарата и др.
Нами показана гетерохрония постнатального морфогенеза нейроцитов гомологичных ганглиев у разных видов животных (рис. 1), различных отделов и ганглиев нервной системы одного вида (рис. 2), однотипных клеток в различных участках кишечника (рис. 3а, в), разнотипных нейроцитов одного ганглия (рис. 3в, г).
В некоторых ганглиях гетерохронно развиваются и однотипные клетки, на что указывают наши и литературные данные о наличии незрелых клеток в интрамулярных ганглиях кишечника, спинальных ганглиях растущих и даже зрелых животных и человека. Гетерохрония морфологии нервных клеток проявляется в объеме их ядра и цитоплазмы, величине ядерно-плазменного отношения, структуре дендритного аппарата и других показателях. Вместе с тем следует отметить, что в некоторых нервных образованиях однотипные нейроциты созревают и развиваются практически синхронно (мотонейроны одного сегмента спинного мозга свиньи и крупного рогатого скота, клетки Пуркинье коры мозжечка собаки и др.).
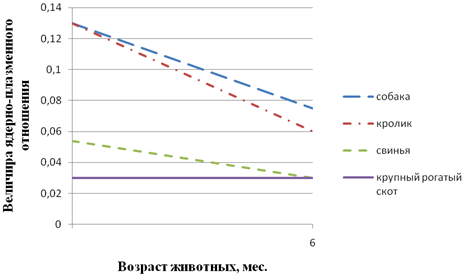
Рис. 1 - Возрастные особенности величины ядерно-плазменного отношения нейроцитов 1-го крестцового спинального ганглия у домашних животных; 1-собаки; 2-кролика; 3-свиньи; 4-крупного рогатого скота
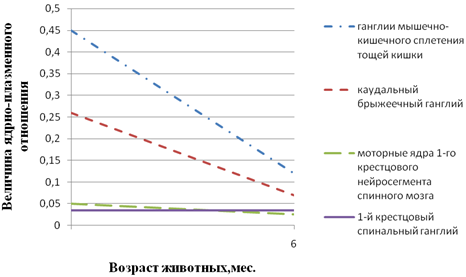
Рис. 2 - Возрастные особенности величины ядерно-плазменного отношения нейроцитов различных ганглиев и ядер свиньи: 1-ганглиев межмышечно-кишечного сплетения тощей кишки; 2-каудального брыжеечного ганглия; 3-моторных ядер 1-го крестцового нейросегмента спинного мозга; 4-1-го крестцового спинального ганглия
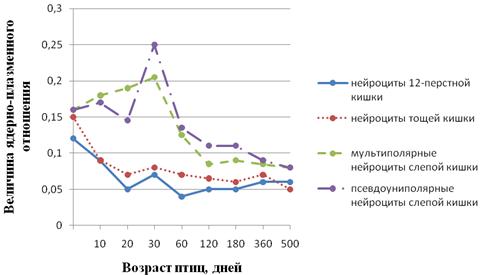
Рис. 3 - Возрастные особенности величины ядерно-плазменного отношения нейроцитов мышечно-кишечного сплетения кур. А-нейроциты 12-перстной кишки; Б-нейроциты тощей кишки; В-псевдоуниполярные нейроциты слепой кишки
Анализ результатов наших исследований показывает, что морфологические преобразования нейроцитов в той или иной степени продолжаются на протяжении сего постнатального онтогенеза животных, однако наибольшим динамизмом отличается в этом отношении начальный этап последнего (рис. 3). Лишь период от наступления зрелости до старости животных характерен относительной стабильности морфологии нейроцитов. Здесь нужно оговориться, что речь идет о достаточно стойких долговременных структурных изменениях нейронов, имея в виду, что экспериментально показаны Ю.Я. Гейнисманом, и значительные кратковременные, реактивные изменения морфологии нервных клеток в связи с изменением интенсивности функций и условий их жизнедеятельности.
На основании анализов результатов накопленного в нашей лаборатории материала мы пришли к заключению, что в основе гетерохронии постнатального (как и пренатального) морфогенеза нейроцитов лежит генетически обусловленная прямая коррелятивная связь между уровнями морфо-функциональной зрелости нервных и иннервируемых структур. Это подтверждается, в частности, данными, приведенными на рис. 1 - 3. Рис. 1 показывает, что у новорожденных телят и поросят, уже способных следовать за матерью, нервные клетки спинальных ганглиев по величине ядерно-плазменного отношения близки таковым половозрелых животных, тогда как у щенков и крольчат, рождающихся почти неспособными к активному передвижению, названный показатель далек от зрелого состояния. Этот же факт противоречит мнению о зависимости уровня зрелости нейроцитов от продолжительности периода внутриутробного развития. У собаки этот период вдвое длиннее, чем у крольчихи (соответственно 62 и 30 дней), а уровень зрелости исследованных нейроцитов одинаков. К аналогичному выводу приводит и анализ данных рис. 2, 3. Из данных рис. 2 следует, что у новорожденных поросят наиболее зрелыми являются мотонейроны спинного мозга, регулирующие деятельность функционально относительно зрелой скелетной мускулатуры. Наименее зрелы нейроциты мышечно-кишечного сплетения тощей кишки, иннервирующие ее компоненты, далекие от зрелого состояния. Рис. 3 свидетельствует об отставании морфогенеза нейроцитов мышечно-кишечного сплетения слепой кишки кур от такового тонкого кишечника. Этот факт согласуется с задержкой начала активной функции слепых кишок, о чем можно судить по отсутствию в них до 30-дневного возраста пищевых масс В.В. Батраков..
Противоречащим изложенному может на первый взгляд показаться гетерохрония постнатального морфогенеза однотипных нейронов одного и того же узла. Действительно, возникает вопрос: чем обусловлена гетерохрония морфологически и функционально однотипных нейроцитов одного спинального ганглия? Тем, по-видимому, обстоятельством, что одни из них осуществляют рецепцию с кожи, другие с аппарата органов движения, третьи внутренностей, то есть структур различного уровня морфо-функциональной зрелости. Аналогичную ситуацию можно предположить и в ганглиях интрамуральных нервных сплетений.
Наличие в интрамуральных ганглиях желудка и кишечника взрослых и даже старых животных значительного количества незрелых нейроцитов послужило некоторым исследователям В.М. Калетина, основанием считать последние «резервом» для восполнения естественной убыли нейронов. Подобная точка зрения вызывает вопрос: почему такого «резерва» до сих пор не обнаружено во многих других ганглиях? Во всяком случае, мы не видели его в моторных ядрах и симпатических ганглиях свиньи, ганглиозном слое коры мозжечка собаки и др. По-видимому, нужно искать другое объяснение этому факту. Возможно, что он связан с образованием в онтогенезе новых нейромоторных и нейросенсорных единиц в органах, структурные элементы которых (гладкая мышечная ткань, эпителий и др.) способны к гиперплазии. Тем более, что компенсация естественной убыли нейронов у старых животных показана за счет гиперплазии дендритов уцелевших клеток Ю.Я. Гейнисмана.
Прямую коррелятивную связь между уровнями морфо-функциональной зрелости нейронов и иннервируемых ими структур мы показываем в качестве одного из основных эндогенных факторов гетерохронного морфогенеза нейроцитов. Вместе с тем мы, как и другие авторы Л.И. Корочкин, и др. отмечаем усиливающиеся в онтогенезе влияния на эти процессы внешних экзогенных факторов, особенно функционального и алиментарного. При этом следует отметить, что названные экзогенные факторы отнюдь не вступают в противоречие с рассмотренным генетическим, а лишь изменяют его подвижность, выраженность и направленность.
Анализ данных, представленных на рис. 3,4 показывает, что наибольшая подвижность морфогенеза свойственна исследованным нейроцитам в раннем постнатальном онтогенезе животных, которому свойственно резкое усиление функциональной активности всех систем организма и, в частности, пищеварительной.
Различная степень выраженности проявлений морфогенеза видна на рис. 3. В третьей декаде после вылупления цыпленка у всех нейроцитов кишечника объем ядра нарастает быстрее объема цитоплазмы, вследствие чего увеличивается ядерно-плазменное отношение. Но особенно значительно увеличивается объем ядер псевдоуниполярных (чувствительных) нейроцитов слепых кишок, что совпадает с началом их активного функционирования.
Направленность идентичных показателей морфогенеза различных нейроцитов в отдельные периоды онтогенеза животных может быть совпадающей и несовпадающей. Примеров первой может служить изменение величины ядерно-плазменного отношения нейроцитов 12-перстной и тощей кишок кур (рис.3а, б). Несовпадающую направленность морфогенетических процессов обнаруживает сравнение изменения объема перикариона нейроцитов интрамуральных (рис.4) и спинальных (рис. 5) узлов теленка на втором месяце после рождения. Если объем первых в этот период значительно уменьшается, то объем вторых последовательно увеличивается. Сам факт разнонаправленности морфогенеза в приведенном примере заслуживает внимания. Следует учесть, что если сенсорная функция нейроцитов спинальных узлов в этот период резких изменений не претерпевает, то в деятельности кишечника наступают значительные изменения, обусловленные переводом телят с молочного на растительный тип кормления. Экспериментально показано Ю.Я. Гейнисманом, что достаточно сильное и длительное раздражение нейроцита приводит к уменьшению объема его ядра и цитоплазмы. Сходное влияние изменения типа кормления на постнатальный морфогенез нейроцитов чревного ганглия нами показано в опыте с ранним (в месячном возрасте) отъемом поросят (рис. 6).
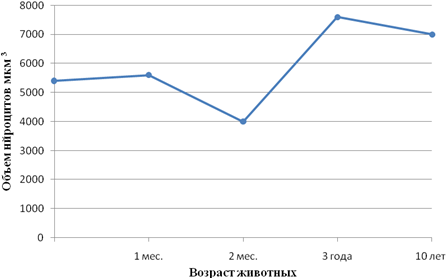
Рис. 4 - Возрастные особенности объема нейроцитов 1-го типа Догеля в ганглиях мышечно-кишечного сплетения слепой кишки крупного рогатого скота
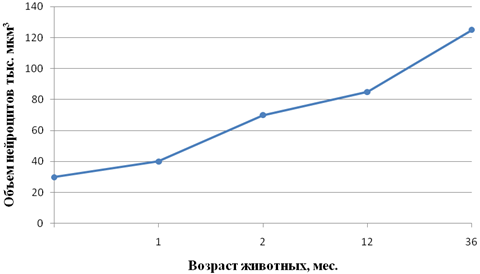
Рис. 5 - Возрастные особенности объема нейроцитов поясничных спинальных ганглиев крупного рогатого скота
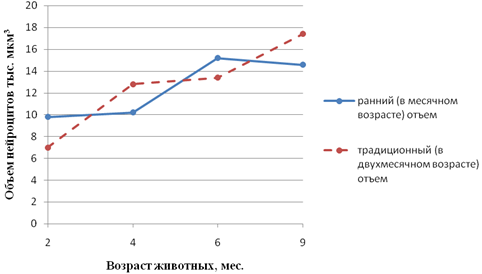
Рис. 6 - Возрастные особенности объема нейроцитов чревного ганглия поросят различных сроков отъема. 1-у поросят раннего (в месячном возрасте) отъема; 2-у поросят традиционнго (в двухмесячном возрасте) отьема
Влияние условий кормления и содержания на постнатальный морфогенез нейроцитов показано нами и на примере интрамулярных нейроцитов кур, содержащихся в клетках в условиях птицефабрики и напольном в условиях личного подсобного хозяйства. Интенсивная технология содержания кур ускоряет постнатальный морфогенез нейроцитов кишечника, но вместе с тем обуславливает значительно более раннее (в 1,5 - летнем возрасте) появление деструктивных явлений (пикнотизация, кариолизис и др.).
Мы далеки от мысли считать свои задачи решенными. Поднятые вопросы нуждаются в силу своей сложности и важности в дальнейшей разработке силами многих исследователей. К тому же рамки одной статьи исключают возможность детального обсуждения даже имеющихся результатов. Вместе с тем изложенное показывает, что при всей сложности, разнообразии и неоднозначности проявлений постнатального морфогенеза нейроцитов он подчинен определенным общим закономерностям, подвержен влиянию не только генетических, но и средовых, экзогенных факторов. Это влияние не может не учитываться в практике ветеринарии и животноводства.
ВЫВОДЫ
На основании полученных нами результатов и с учетом данных других исследователей мы считаем целесообразным выделить в постнатальном морфогенезе нейроцитов млекопитающих и птиц несколько периодов, отличающихся друг от друга подвижностью, выраженностью и направленностью формообразовательных процессов.
- Ранний постнатальный период. В зависимости от продолжительности жизни вида длится от нескольких месяцев (домашние млекопитающие и птицы) до нескольких лет (человек). Характерными особенностями этого периода являются большая подвижность, выраженность морфогенеза нейроцитов, прогрессивная (ассимилятивная) его направленность.
- Период полового созревания. Продолжается от начала полового созревания до наступления зрелости тела. Характерен умеренной подвижностью, выраженностью и прогрессивной направленностью морфогенеза нейроцитов, уравновешенностью прогрессивной (ассимилятивной) и регрессивной (диссимилятивной) их направленности.
- Старческий период. Его началом можно считать прекращение репродуктивной функции у самок и соответствующий возраст самцов. Для него характерны малая подвижность и выраженность морфогенеза нейроцитов, преобладание регрессивной (диссимилятивной) его направленности.
Список литературы / References
- Гистогенез дистального ганглия блуждающего нерва свиньи /Симанова Н.Г., Хохлова С.Н.//В сборнике: Аграрная наука и образование на современном этапе развития: опыт, проблемы и пути их решения. ФГОУ ВПО, Ульяновская государственная сельскохозяйственная академия; 2009.- С. 102-104.
- Использование музейных экспонатов по морфологии в учебном процессе / Н.Г. Симанова, С.Н. Хохлова, А.Н. Фасахутдинова, Т.Г. Скрипник //В сборнике: Инновационные технологии в высшем профессиональном образовании материалы.- Ульяновская ГСХА.- 2010.- С. 160-163.
- Импрегнация нервных структур мозжечка крысы по методу Бильшовского-Грос / И.Д.Ипастова, Н.П.Перфильева, С.Н.Хохлова, С.Г.Писалёва, Н.Г.Симанова // Материалы международной научно-практической конференции.- Саранск, 2015.- С.66-69.
- Использование музейных экспонатов по морфологии в учебном процессе / Симанова Н.Г., Скрипник Т.Г., Хохлова С.Н., Фасахутдинова А.Н.// В сборнике: Инновационные технологии в высшем профессиональном образовании материалы Научно-методической конференции профессорско-преподавательского состава академии. 2010.- С. 160-163.
- Морфогенез нервной системы домашних животных /Симанова Н.Г., Хохлова С.Н., Фасахутдинова А.Н. Немецкая Национальная Библиотека. Saarbrucken, 2014.
- Морфогенез спинальных ганглиев и спинного мозга кролика /А.Н.Фасахутдинова, Н.Г. Симанова, С.Н. Хохлова // Сборник статей международной научно-практической конференции.- Уфа, Аэтерна, 2015.- С. 32-42.
- Симанова, Н.Г. Возрастные особенности миелоархитектоники шейного отдела блуждающего нерва свиньи и собаки / Н.Г. Симанова, Т.Г. Скрипник // Вестник Ульяновской государственной сельскохозяйственной академии.- 2007.- № 1.- С. 62-64.
- Симанова, Н.Г. Возрастные особенности строения стенки сфинктеров пищеварительной трубки у собак / Н.Г. Симанова // Материалы международной научно-практической конференции, 2003.- С. 71-72.
- Учебная практика по анатомии домашних животных / Н.А. Жеребцов, Н.Г. Симанова, С.Н. Хохлова, А.Н. Фасахутдинова, В.М. Елин // Методические указания для студентов 1 курса по специальности "Ветеринария" / Ульяновская ГСХА, 2004. – 45 с.
- Хохлова, С.Н Возрастные изменения морфологии нервных узлов половой системы самок домашних животных/ С.Н. Хохлова, Н.Г. Симанова, М.А. Богданова, А.А. Степочкин, А.Н. Фасахутдинова //Международный научно-исследовательский журнал. – Екатеринбург, - № 4 (46). - Ч.5 . - С. 65-68.
Список литературы на английском языке / References in English
- Gistogenez distalnogo gangliya bluzhdayushchego nerva svinyi [Histogenesis of Distal Nerve Center of Vagus Nerve of Pig] / Simanova N.G., Khokhlova S.N. // Sbornik: Agrarnaya nauka I obrazovaniye na sovremennom etape razvitiya: opyt, problemy i puti ikh resheniya проблемы [Collected papers: Agrarian science and education at the present stage of development: experience, problems and ways to solve them] FSEI HPE, Ulyanovsk State Agricultural Academy; 2009. – P. 102-104. [In Russian]
- Ispolzovaniye muzeynykh eksponatov po morfologii v uchebnom protsesee [Use of Museum Exhibits on Morphology in Educational Process] / N.G. Simanova, S.N. Khokhlova, A.N. Fasakhutdinova, T.G. Skripnik // Sbornik: Innovatsionniye tekhnologii v vysshem professionalnom obrazovaniyi materialy [Collected papers: Innovative technologies in higher professional education materials] - USAA.- 2010. – P. 160-163. [In Russian]
- Impregnatsiya nervnykh struktur mozzhechka krysy po metodu Bilshkovskogo-Gros [Impregnation of Nerve Structures of Parencephalon of Rat According to Bilshovsky-Gros Method] / I.D. Ipastova, N.P. Perfilyeva, S.N. Khokhlova, S.G. Pisaleva, N.G. Simanova // Materialy mezhdunarodnoy nauchno-prakticheskoy konferentsii [Materials of the International Scientific and Practical Conference] - Saransk, 2015. – P.66-69 [In Russian]
- Ispolzovaniye muzeynykh eksponatov po morfologii v uchebnom protsesse [Use of Museum Exhibits on Morphology in Educational Process] / Simanova N.G., Skripnik T.G., Khokhlova S.N., Fasakhutdinova A.N. // Sbornik: Innovatsionniye tekhnologii v vysshem professionalnom obrazovanii, materialy Nauchno-metodicheskoy konferentsii profesorssko-prepodavatelskogo sostava akademiyi [Collected papers: Innovative technologies in higher professional education, materials of the Scientific and Methodological Conference of Faculty members of the Academy]. 2010. – P. 160-163. [In Russian]
- Morfogenez nervnoy sistemy domashnikh zhivotnykh [Morphogenesis of Nervous System of Domestic Animals] / Simanova N.G., Khokhlova S.N., Fasakhutdinova A.N. German National Library. Saarbrucken, 2014. [In Russian]
- Morfogenez spinalnykh gangliev i spinnogo mozga krolika [Morphogenesis of Spinal Nerve Center and Spinal Cord of Rabbit] / A.N. Fasakhutdinova, N.G. Simanova, S.N. Khokhlova // Sbornik statey mezhdunarodnoy nauchno-prakticheskoy konferentsii [Collected papers: The international scientific and practical conference] - Ufa, Aeterna, 2015.- P. 32-42. [In Russian]
- Simanova, N.G. Vozrastniye osobennosti mieloarkhitoniki sheynogo otdela bluzhdayushchego nerva svinyi i sobaki [Age Features of Myeloarchitectonics of Cervical Region of Vagus Nerve of Pig and Dog] / N.G. Simanova, T.G. Skripnik // Vestnik Uliyanovskoy gosudarstvennoy selskokhoziaystvennoy akademii [Bulletin of the Ulyanovsk State Agricultural Academy]. - 2007. – No. 1. – P. 62-64. [In Russian]
- Simanova, N.G. Vosrastniye osobennosti stroyeniya stenki sfinkterov pishchevaritelnoy trubki u sobak [Age Features of the Sphincter Wall Structure of Digestive Duct of Dog] / Simanova N.G. // Materialy mezhdunarodnoy nauchno-prakticheskoy konferentsii [Proceedings of the International Scientific and Practical Conference] 2003. – P. 71-72. [In Russian]
- Uchebnaya praktika po anatomii domashnikh zhivotnykh [Training on Anatomy of Domestic Animals] / N.A. Zherebtsov, N.G. Simanova, S.N. Khokhlova, A.N. Fasakhutdinova, V.M. Elin // Metodicheskiye ukazaniya dlia studentov 1 kursa po spetsialnosti "Veterinariya" [Methodological instructions for 1st year students on the specialty "Veterinary"] / Ulyanovsk State Agricultural Academy, 2004. – 45 p. [In Russian]
- Khokhlova, S.N. Vozrastniye izmeneniya morfologii nervnykh uzlov polovoy sistemy samok domashnikh zhivotnykh [Age-related Changes in Morphology of Nerve Knots of Reproductive System of Female Domestic Animals] / S.N. Khokhlova, N.G. Simanova M.A. Bogdanova, A.A. Stepochkin, A.N. Fasakhutdinova // Mezhdunarodniy nauchno-issledovatelskiy zhurnal [International Scientific and Research Journal] - Ekaterinburg, 2016. - No 4 (46). - P.5. – P. 65-68. [In Russian]