Магнитно-резонансная томография в диагностике функциональных изменений головного мозга У ДЕВОЧЕК-подростков С НОРМАЛЬНЫМ МЕНСТРУАЛЬНЫМ ЦИКЛОМ
Магнитно-резонансная томография в диагностике функциональных изменений головного мозга У ДЕВОЧЕК-подростков С НОРМАЛЬНЫМ МЕНСТРУАЛЬНЫМ ЦИКЛОМ
Аннотация
Цель: определение функциональных изменений головного мозга во время нормального менструального цикла у девочек-подростков с помощью функциональной магнитно-резонансной томографии (фМРТ) в состоянии покоя.
Материалы и методы: анализ данных фМРТ головного мозга в состоянии покоя 39 относительно здоровых девочек-подростков в возрасте от 13 до 17 лет (средний возраст 16,3 ±0,7) с регулярным менструальным циклом. Статистический анализ и оценка полученных данных фМРТ покоя проводили при применении плагина CONN v.22а – Functional connectivity toolbox, на базе MATLAB.
Функциональная МРТ покоя позволяет оценить базовую активность мозга в отсутствии определенных сенсорных или когнитивных стимулов и визуализировать рабочие сети, включающие различные участки головного мозга, демонстрирующие синхронные изменения BOLD-сигнала в состоянии покоя. Существует ряд закономерностей изменения BOLD-сигнала, которые можно наблюдать в различные фазы менструального цикла.
Использование фМРТ может помочь в понимании глубоких нейрофизиологических процессов, происходящих в организме девочки-подростка во время нормального менструального цикла, а также открыть новые возможности в диагностике функциональных нарушений головного мозга у девочек-подростков с нарушениями менструального цикла.
1. Введение
Репродуктивное здоровье детей и подростков в системе общих ценностей формирования здоровья населения занимает одно из ключевых мест, обеспечивая продолжение роста населения страны и человеческого рода в целом.
Индикатором репродуктивного здоровья женщины является менструация. Менструальный цикл (МЦ) (от латинского menstruus – месячный, ежемесячный) – это циклические изменения в организме женщины, преимущественно в звеньях репродуктивной системы, повторяющиеся через определённые промежутки времени и проявляющиеся регулярными маточными кровотечениями – менструациями (menses). Нормальный МЦ является результатом нейрогормональных взаимоотношений между ЦНС, гипоталамусом, гипофизом, яичниками и маткой. Регулярный менструальный цикл устанавливается в течение года после первой менструации (menarche) и сохраняется до 45-52 лет. Для среднеевропейской зоны возраст менархе составляет 12-13 лет , , , .
Изучение влияния гормонов на головной мозг во время менструального цикла в настоящее время является популярной областью для исследований при помощи методов нейровизуализации. Нейровизуализация стала предпочтительным инструментом для установления гормональных эффектов в головном мозге
, .С помощью функциональной магнитно-резонансной томографии (фМРТ) неоднократно проводились исследования головного мозга в разные фазы менструального цикла, в которых доказывалось, что колебания уровня эндогенных половых стероидов влияют на структуру и функциональную организацию мозга
, .Однако большинство исследований проводятся у взрослых женщин, упуская из внимания девочек-подростков, у которых менструальный цикл может иметь свои визуализационные особенности в связи с несовершенством регуляции работы гипатоламо-гипофизарно-яичниковой системы.
Именно поэтому целью исследования являлось определение функциональных изменений головного мозга во время регулярного менструального цикла у девочек-подростков путем проведения функциональной МРТ в состоянии покоя.
2. Методы и принципы исследования
Проведен анализ данных фМРТ головного мозга в покое 39 здоровых девочек-подростков в возрасте от 13 до 17 лет (средний возраст 16,3 ±0,7) с регулярным менструальным циклом. Подтверждение фазы менструального цикла проводилось с помощью индивидуального календаря менструации, представленного пациентками.
МР-исследование проводили двукратно в фолликулярную (с 5-15 день) и лютеиновую (с 15-25 день) фазы менструального цикла соответственно на 1,5 Тл томографе с применением стандартных импульсных последовательностей (Т1-WI, Т2-WI, TIRM толщина среза – 3,5 мм; MPRAGE толщина среза – 1,2 мм; DWI), последовательностей с прицелом на гипоталамо-гипофизарную зону (толщина среза – 3 мм) и последовательности BOLD для оценки функциональной коннективности головного мозга в состоянии покоя.
Статистический анализ и оценка полученных данных фМРТп проводили при применении плагина CONN v.22а – Functional connectivity toolbox, на базе MATLAB, который позволяет выявить взаимосвязи между различными структурами головного мозга и подтвердить их зоны активации с помощью картирования.
В исследовании использовали анализ оценки низкоуровневой корреляции (ROI-to-ROI – анализ данных на основе выбора зоны интереса для оценки функциональной коннективности с другими зонами) и групповой анализ независимых компонентов (Independent component analysis, ICA), пороговое значение pFDR <0,05.
Предварительная обработка: Функциональные и анатомические данные были предварительно обработаны с использованием гибкого конвейера предварительной обработки, включая перестройку с коррекцией взаимодействия искажений чувствительности, коррекцию времени среза, обнаружение выбросов, прямую сегментацию и нормализацию MNI-пространства, а также сглаживание. Функциональные данные были перестроены с использованием процедуры SPM realign & unwarp где все сканы были привязаны к эталонному изображению (первое сканирование первого сеанса) с использованием метода наименьших квадратов и преобразования с 6 параметрами, и повторно выбраны с использованием b-сплайновой интерполяции для коррекции движения и взаимодействия магнитной восприимчивости. Временное рассогласование между различными фрагментами функциональных данных (полученных в порядке возрастания) было исправлено с помощью процедуры SPM slice-timing correction (STC) , используя временную интерполяцию sinc для повторной выборки каждого фрагмента временных рядов до общего среднего времени сбора. Потенциальные снимки с выбросами были идентифицированы с использованием ART как снимки со смещением по кадру более 0,9 мм или глобальными изменениями сигнала выше 5 стандартных отклонений и эталонное изображение было вычислено для каждого объекта путем усреднения всех снимков, исключая выбросы. Функциональные и анатомические данные были нормализованы в стандартном пространстве MNI, сегментированы на классы тканей серого вещества, белого вещества и цереброспинальной жидкости и повторно отобраны до изотропных вокселов размером 2 мм после процедуры прямой нормализации с использованием унифицированного алгоритма сегментации и нормализации SPM с шаблоном карты вероятности ткани IXI-549 по умолчанию. Наконец, функциональные данные были сглажены с использованием пространственной свертки с гауссовым ядром, равным половине максимальной ширины 8 мм (FWHM).
Шумоподавление: кроме того, функциональные данные были шумоизолированы с использованием стандартного конвейера шумоподавления, включая регрессию потенциальных смешивающих эффектов, характеризуемых временными рядами белого вещества (5 компонентов шума CompCor), временными рядами CSF (5 компонентов шума CompCor), параметрами движения и их производными первого порядка (12 факторов) сканирование выбросов (менее 7 факторов), эффекты сеанса и задачи и их производные первого порядка (6 факторов), а также линейные тренды (2 фактора) в рамках каждого функционального запуска, затем следует полосовая частотная фильтрация временных рядов в диапазоне от 0,008 Гц до 0,09 Гц. CompCor компоненты шума в белом веществе и ликворе оценивались путем вычисления среднего сигнала BOLD, а также наибольших основных компонентов, ортогональных среднему значению BOLD, параметров движения и сканов выбросов в размытых масках сегментации каждого субъекта. Исходя из количества шумовых терминов, включенных в эту стратегию шумоподавления, эффективные степени свободы сигнала BOLD после шумоподавления варьировали от 125 до 132,8 (в среднем 131,3) у всех испытуемых.
3. Основные результаты
При проведении анализа на основе выбора зоны интереса для оценки функциональной коннективности с другими зонами (ROI-to-ROI – при условии Difference Follicular> Luteal) было установлено, что в фолликулярную фазу менструального цикла коннективность была повышена между (табл.1., рис 1.):
- височно-затылочной частью средней височной извилины слева и передним отделом средней височной извилины слева;
- нижней лобной извилиной слева и нижней лобной извилиной справа;
- задним отделом средней височной извилины справа и дорсальной сетью внимания (внутритеменной бороздой слева);
И в лютеиновую фазу менструального цикла между (табл.1., рис 1.):
- верхним отделом латеральной затылочной коры справа и верхним отделом латеральной затылочной коры слева;
- верхним отделом латеральной затылочной коры справа и сетью покоя по умолчанию слева;
- сетью покоя по умолчанию справа и сетью покоя по умолчанию слева;
- зрительной корой справа и сетью покоя по умолчанию справа;
- нижней лобной извилиной слева и треугольной частью нижней лобной извилины справа;
- зрительной корой справа и верхним отделом латеральной затылочной коры слева.
Таблица 1 - Области повышенной и сниженной (с отрицательной Т) коннективности рабочих сетей в фолликулярную фазу по сравнению с лютеиновой
Analysis Unit | Statistic T (7) | p-unc | p-FDR |
Lateral Occipital Cortex, superior division Right (sLOC r) - Lateral Occipital Cortex, superior division Left (sLOC l) | -21,20 | 0,000000 | 0,001748 |
Lateral Occipital Cortex, superior division Right - networks Default Mode Lateral Parietal Left (Default Mode LP l) | -13,91 | 0,000002 | 0,015666 |
Middle Temporal Gyrus, temporooccipital part Left (pMTG l) - Middle Temporal Gyrus, anterior division Left (aMTG l) | 11,94 | 0,000007 | 0,029235 |
Networks Default Mode Lateral Parietal Right - networks. Default Mode Lateral Parietal Left | -10,86 | 0,000012 | 0,041327 |
Networks Visual Lateral Right - networks Default Mode Lateral Parietal Right | -9,92 | 0,000023 | 0,049552 |
networks Language Inferior Frontal Gyrus Left - Inferior Frontal Gyrus, pars triangularis Right | 9,73 | 0,000026 | 0,049552 |
networks Visual Lateral Left - Occipital Pole Right (OP r) | -9,65 | 0,000027 | 0,049552 |
networks Default Mode. Lateral parietal Right - Lateral Occipital Cortex, superior division Left (sLOC l) | -9,46 | 0,000031 | 0,049552 |
Middle Temporal Gyrus, posterior division Right – networks Dorsal Attention. Intraparietal sulcus Left (IPS l) | 9,35 | 0,000033 | 0,0495 |
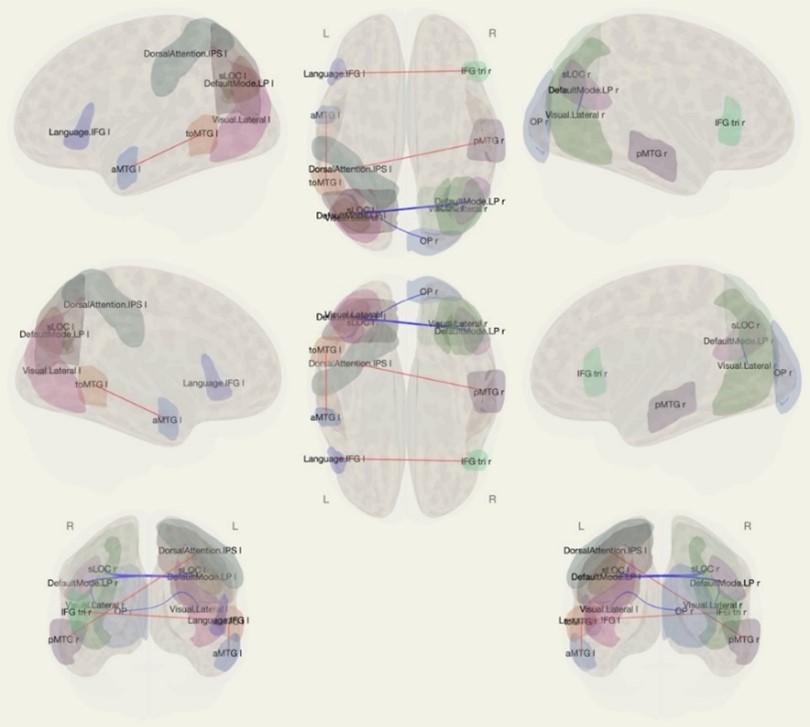
Рисунок 1 - Результаты анализа ROI-to-ROI
Примечание: аббревиатуры, представленные на изображении расшифрованы в таблице; графическое представление результатов, полученных субтракцией функциональных метрик в фолликулярную фазу менструального цикла из данных в лютеиновую фазу менструального цикла; красным отмечены положительные, а синим отрицательные функциональные связи между рабочими сетями головного мозга (р <0,05)
Кластер 1 представлен передним и задним отделами надмаргинальной извилины слева, теменной корой слева, постцентральной и угловой извилинами слева.
Кластер 2 представлен передним и задним отделами надмаргинальной извилины справа, прецентральной и постцентральной извилинами справа.
Кластер 3 представлен средней лобной извилиной слева и медиальной лобной извилиной слева (рис. 2).
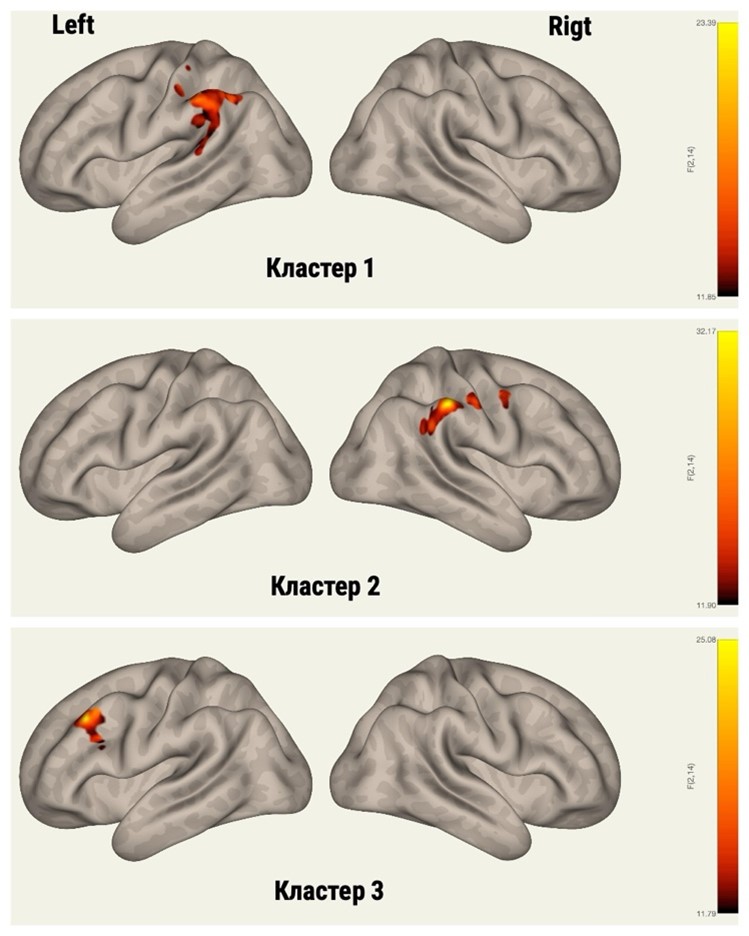
Рисунок 2 - Кластеры головного мозга входящие в состав визуальной сети покоя ()
Примечание: ICA 18, 20; кластер 1 - 296 (49%) вокселей покрывающие 31% переднего отдела супрамаргинальной извилины слева, 193 (32%) вокселя покрывающие 18% заднего отдела супрамаргинальной извилины слева, 37 (6%) вокселей покрывающие 7% теменной коры головного мозга слева, 21 (3%) воксель покрывающие 1% постцентральной извилины слева, 20 (3%) вокселей покрывающие 2% угловой извилины слева, 6 (1%) вокселей покрывающие 1% височного планума слева, 5 вокселей (1%) покрывающие 1% заднего отдела верхней височной извилины слева; кластер 2 - 143 (32%) вокселя покрывающие 18% переднего отдела супрамаргинальной извилины справа, 98 (22%) вокселей покрывающие 3% постцентральной извилины справа, 96 (21%) вокселей покрывающие 8% заднего отдела супрамаргинальной извилины справа; кластер 3 - 207 (87%) вокселей покрывающие 7% медиальной лобной извилины слева, 4 (2%) вокселя покрывающие 1% оперкулярной части нижней лобной извилины слева
4. Обсуждение
В результате проведенного исследования следует, что одними из наиболее функционально активных сетей головного мозга в фолликулярную и лютеиновую фазы менструального цикла являются: рабочая сеть по умолчанию (Default Mode Network – DMN, сеть «холостого хода» или пассивного режима работы мозга – СПРРМ), зрительная сеть (Visual Network – VN) и дорсальная сеть внимания (Dorsal Attention Network – DAN). Также, исходя из полученных данных, становится понятно, что регуляция менструального цикла осуществляется, в том числе, и с помощью межполушарных взаимодействий. Чтобы глубже понять, какие функциональные процессы головного мозга в состоянии покоя тесно связаны с менструальным циклом, стоит сказать о каждой из вышеописанных сетей более подробно.
Сеть пассивного режима работы мозга – это крупномасштабная сеть мозга, которая включает в себя несколько высокоуровневых когнитивных областей, таких как медиальная префронтальная кора (medial Prefrontal Cortex – mPFC), задняя часть поясной коры (Posterior Cingulyte Cortex – PCC) и задние теменные области (Posterior Temporal Lobe – PTL). DMN в основном известна как сеть, регионы которой демонстрируют активность в покое и деактивируются во время когнитивных целевых задач . Тем не менее DMN также может быть активирована во время высокоуровневых социальных когнитивных задач в подростковом возрасте . Хотя точная функция DMN все еще не полностью понятна, считается, что DMN поддерживает самореферентную деятельность и состоит из регионов, участвующих в различных когнитивных функциях. Регионы в DMN мало связаны у детей в возрасте от 7 до 9 лет, но сильно связаны у взрослых, что указывает на то, что детство и подростковый возраст являются критическими периодами для развития этой ключевой мозговой сети. Особое значение имеет отсутствие передних и задних связей в раннем возрасте. Имеющиеся данные свидетельствуют о том, что DMN способствует умственному исследованию социального и эмоционального содержания. Активность DMN увеличивается во время перспективного восприятия желаний, убеждений и намерений (т.е. теории разума), при запоминании прошлого (например, автобиографическая память) и при планировании будущего. Эти функции предполагают, что DMN может способствовать адаптивному поведению, позволяя создавать, воспроизводить и исследовать сценарии в уме, как для размышлений о прошлых событиях, так и для выведения ожиданий в отношении будущего. Снижение активности в DMN во время усиленной когнитивной обработки может отражать необходимость снижения самореферентной активности мозга, чтобы сосредоточиться на внешней поставленной задаче. Так, например, чрезмерная активность DMN может помочь объяснить общие симптомы при депрессии. Депрессия характеризуется повышенным размышлением или рецидивирующим, рефлексивным и неконтролируемым фокусом на депрессивном настроении и его причинах и последствиях. Депрессия у взрослых среднего возраста неоднократно ассоциировалась с повышенной активностью в DMN что, в свою очередь, связано с отрицательным размышлением . В настоящее время существуют исследования подтверждающие функциональные изменения головного мозга во время предменструального дисфорического расстройства , , а также различные паттерны нарушения коннективности мозга, связанные с депрессией и нарушением менструального цикла , , .
Зрительная сеть включает в себя боковую (или вторичную) зрительную сеть, медиальную (или первичную) визуальную сеть и затылочную зрительную сеть (пространственную). Боковая зрительная сеть состоит из перистриатной области, боковой и верхней затылочной извилины. Медиальная визуальная сеть включает в себя полосатую и парастриатную части. Затылочная визуальная сеть, участвующая в визуальной обработке более высокого уровня, связанной с орфографией и скрытым чтением, которая включает в себя в основном затылочный полюс (затылочно-теменной узел), медиальные отделы предклинья, задние отделы поясной извилины и лобные доли. Эта сеть ответственна за пространственное восприятие зрительной информации. Визуальная сеть участвует в обнаружении потенциально значимых стимулов для организма. Эти визуальные стимулы могут повысить или понизить значимость болезненного фактора, что приводит к увеличению или уменьшению боли. Например, наблюдение за тем, как игла колет руку другого человека, создает субъективное ощущение дискомфорта и может повышать степень боли при проведении подобной манипуляции с наблюдающим. Известно, что колебания половых гормонов во время менструального цикла приводят к значимым изменения в ощущении боли женщинами, а также значительно влияют на степень восприятия боли и её различия между мужчинами и женщинами .
Дорсальная сеть внимания – это последовательная, двусторонняя сеть для постоянного удержания внимания. Поскольку эффективные, целенаправленные процессы DAN являются ключевыми элементами познания, сеть тесно связана со многими другими функциональными областями мозга. Существует два режима обработки внимания: внимание снизу вверх и сверху вниз. Обработка сверху вниз, основана на пространственном внимании и позволяет выбирать, где сосредоточить внимание на основе ожиданий и сознательных целей. Обработка снизу вверх – это эффективный сдвиг внимания в ответ на предсознательный стимул, такой как движение, неожиданные шумовые или визуальные изменения. Сеть вентрального внимания мозга (Ventral Attention Network – VAN) отвечает за сдвиг снизу вверх. Когда внимание смещается, сеть последовательно выделяет элементы в визуальном поле по мере того, как происходит последовательная обработка. Чтобы обеспечить гармоничное смещение внимания, DAN работает не в одиночку. Другие мозговые сети также помогают в реагировании на стимулы и управление фокусом. Сеть вентрального внимания переключает внимание в ответ на неожиданные стимулы, такие как шок, пугающие события или «странные» явления. Эта реакция переориентации происходит в одностороннем порядке в правом полушарии с функциональными областями в височно-теменной коре и нижней лобной коре. В то время как VAN в одностороннем порядке расположена в правом полушарии, DAN является двусторонней сетью. Это демонстрирует сильную связь между областями в латеральной затылочной доле, прецентральной борозде, самой дорсальной части верхней лобной борозды, которая считается лобными полями глаз (Frontal Eye Fields – FEF), вентральной премоторной корой, верхней теменной долей, внутритеменной бороздой и чувствительной к движению средней височной областью .
Несмотря на разделение сетей по степени их вовлеченности в тот или иной процесс, все они работают в тесной взаимосвязи и не находятся в вакууме, что позволяет контролировать/изменять/влиять на различные физиологические процессы в головном мозге и организме человека в целом. Менструальный цикл является сложным механизмом, вероятно, регулирующимся не только эндогенными гормонами и нейромедиаторами, но ещё и внутренними сетями головного мозга. Более детальные механизмы, паттерны изменения функциональной коннективности, а также структурные изменения головного мозга во время нормального и нарушенного менструального цикла у подростков ещё предстоит изучить.
5. Заключение
Таким образом, физиологический процесс менструального цикла, который характеризуется сменой гормонального статуса, находит свое отражение в функциональной коннективности различных областей головного мозга.
Это приводит к колебаниям психоэмоционального состояния и когнитивных функций за счет доминирующего вклада таких структур головного мозга, как латеральная теменная кора, компоненты визуальной нейросети, являющимися структурами затылочной доли головного мозга и их обширными связями, а также средняя височная извилина, которая входит в состав дорзальной сети внимания.
Все описанные структуры обуславливают клинические колебания физиологического менструального цикла здоровых подростков.