РИТМИЧНОСТЬ НАРАСТАНИЯ ГОДИЧНЫХ ПОБЕГОВ ИВЫ ТРЕХТЫЧИНКОВОЙ (SALIX TRIANDRA L.) КАК ФАКТОР ОПТИМИЗАЦИИ ПЕСТИЦИДНОЙ НАГРУЗКИ В НАСАЖДЕНИЯХ ИНТЕНСИВНОГО ТИПА
РИТМИЧНОСТЬ НАРАСТАНИЯ ГОДИЧНЫХ ПОБЕГОВ ИВЫ ТРЕХТЫЧИНКОВОЙ (SALIX TRIANDRA L.) КАК ФАКТОР ОПТИМИЗАЦИИ ПЕСТИЦИДНОЙ НАГРУЗКИ В НАСАЖДЕНИЯХ ИНТЕНСИВНОГО ТИПА
Научная статья
Афонин А.А.1, *, Гостева Е.С.2, Ларина Е.С.3
1 ORCID: 0000-0002-9392-2527;
1, 2, 3 Брянский государственный университет имени академика И.Г. Петровского. Брянск, Россия
* Корреспондирующий автор (afonin.salix[at]gmail.com)
АннотацияДана общая характеристика ивы трехтычинковой (Salix triandra L.): таксономия, цитогенетика, практическое значение как источника прута для плетения. Проанализирована возможность генетической защиты насаждений ивы трехтычинковой от патогенов и вредителей. Обоснована необходимость применения пестицидов с пролонгированным действием на прутяных плантациях интенсивного типа. Для снижения пестицидной нагрузки предложено планировать агролесотехнические мероприятия с учетом ритмов развития годичных побегов. В модельной инбредной популяции сеянцев ивы трехтычинковой третьей генерации выявлены закономерности сезонной цикличности изменения длины междоузлий с использованием амплитудно-фазового анализа. Доказано, что сезонные тренды динамики длины междоузлий зависят от индивидуальных особенностей сеянцев и обусловлены взаимодействием низших несинхронизированных гармоник с периодом колебаний 48…144 (288) сут. Цикличность сезонной динамики длины междоузлия, не зависящая от индивидуальных особенностей сеянцев, в большинстве случаев определяется, прежде всего, синхронизированными гармониками с периодом колебаний 36 сут и модулирующими гармониками с периодом колебаний 24 сут. Высшие гармоники с периодом колебаний 10…21 сут оказывают незначительное модулирующее влияние на сезонную динамику длины междоузлия. Для оптимизации пестицидной нагрузки предложено проводить трехкратную обработку насаждений с периодичностью порядка 36 сут.
Ключевые слова: Salix triandra, генетическая защита, пестицидная нагрузка, сезонная динамика, динамика развития.
GROWTH RHYTMICITY OF ANNUAL TWIGS OF ALMOND-LEAVED WILLOW (SALIX TRIANDRA L.) AS PESTICID LOAD OPTIMIZATION FACTOR IN INTENSIVE TYPE PLANTING
Research article
Afonin A.A.1, *, Gosteva E.S.2, Larina E.S.3
1 ORCID: 0000-0002-9392-2527;
1, 2, 3 I. Petrovsky Bryansk State University, Bryansk, Russia
* Corresponding author (afonin.salix[at]gmail.com)
AbstractThe authors present general characteristics of almond-leaved willow (Salix triandra L.): taxonomy, cytogenetics, practical importance as a source of twig for weaving. The authors analyzed the possibility of genetic protection of almond-leaved willow twigs from pathogens and pests in the paper. The necessity of the use of pesticides with a prolonged effect on the twig plantations of the intensive type has been substantiated. In order to reduce the pesticide load, they proposed to plan agroforestry measures which take into account the rhythms of development of annual twigs. In the model inbred population of almond-leaved willow seedlings of the third generation, the patterns of seasonal cyclicity of changes in the length of internodes were revealed with the use of amplitude-phase analysis. It is proved that seasonal trends in the dynamics of the length of internodes depend on the individual characteristics of seedlings and are due to the interaction of the lower unsynchronized harmonics with the oscillation period of 48...144 (288) days. The cyclical nature of the seasonal dynamics of the length of the internode, which does not depend on the individual characteristics of seedlings, in most cases is determined primarily by synchronized harmonics with a period of oscillations of 36 days and modulating harmonics with a period of oscillations of 24 days. Higher harmonics with a period of oscillations of 10...21 days have a slight modulating effect on the seasonal dynamics of the length of the internode. To optimize the pesticide load, they proposed to carry out triple treatment of plantations with a frequency of about 36 days.
Keywords: Salix triandra, genetic protection, pesticidal load, seasonal dynamics, development dynamics.
Ива трехтычинковая – Salix triandra L. 1753 (S. amygdalina L. 1753) – широко известный представитель ив Восточной Европы [1, С. 107], включая среднюю полосу России [2, С. 167]. Биология рассматриваемого вида достаточно хорошо изучена. В системе рода Salix L. 1753 ива трехтычинковая – типовой вид секции Amygdalinae Koch 1837 подрода Salix (=Amerina, =Armena) Dum. 1825 [1, С. 103, 104]. Данные молекулярной филогенетики свидетельствуют об особом таксономическом статусе этого вида [3, С. 947]. С цитогенетической точки зрения, S. triandra – диплоид (2n = 2х при x = 19, 22) [4, С. 364], однако в некоторых выборках обнаруживаются триплоиды и тетраплоиды [5, С. 271], [6, С. 622], [7, С. 56]. Столь высокие хромосомные числа указывают на возможность гибридогенного генезиса цитогеномов современных видов Salix [5, С. 282]. Возможность аллополиплоидного происхождения S. triandra была подтверждена нами ранее [8, С. 33].
С практической точки зрения, ива трехтычинковая – источник древесной биомассы для нужд биоэнергетики [9, С. 102] и может использоваться для создания энергетических плантаций [10, С. 43], [11, С. 158]. Однако, в первую очередь, ива трехтычинковая – источник высококачественного прута, пригодного для плетения разнообразных изделий [12, С. 8]. Один из основателей отечественной школы саликологов Э.Э. Керн указывал, что «S. amygdalina дает как зеленый, так и белый, как толстый, так равно и тонкий прут, пригодный и для грубых и для тонких изделий; легко расщепляется и выстругивается в ленты; древесина ее отличается особенною вязкостью и белизною» [13, С. 9,10]. В настоящее время известно около 70 культиваров ивы трехтычинковой, предназначенных для выращивания на прутяных плантациях [14, С. 145].
Благодаря исключительной неприхотливости ивы трехтычинковой экстенсивные плантации на основе этого вида могут закладываться на бедных почвах, малопригодных для выращивания большинства сельскохозяйственных культур [12, С. 8], [15, С. 1], [16, С. 324]. Для повышения продуктивности плантаций и получения высококачественного прута необходимо обеспечить защиту растений от патогенов и вредителей [17, С. 57]. Значительный ущерб ивовым плантациям наносят грибы Melampsora spp., вызывающие ржавчину листьев [18, С. 915]. Генетическая резистентность ив к этому патогену может быть обеспечена селекцией, нацеленной на усиление экспрессии соответствующих генов [19, С. 321]. Листья ив поражаются также различными фитофагами, например, листоедами Phratora (Phyllodecta) vulgatissima [20, С. 289]. Генетическая защита от этого дефолирующего вредителя основана на использовании гибридных клонов, в листьях которых повышено содержание фенолов [21, С. 516]. Продуктивность ивовых плантаций снижается и при воздействии других вредителей: тлей, клешей и т.п. Для снижения ущерба от них также разрабатывается генетическая защита, основанная на использовании гибридов [22, С. 619]. Однако качество прута снижается, в первую очередь, при поражении побегов скрытыми вредителями – побеговыми галлицами рода Dasineura (=Rhabdophaga), в частности, D. marginemtorquens, D. auritae, D. ingeris, D. rosaria [23, С. 163].
Таким образом, создание генетической защиты [24, С. 13], основанной на создании коллекций резистентных культиваров ив (включая гибридные клоны), представляет собой перспективное направление селекции, позволяющее в будущем снизить ущерб от патогенов и вредителей на ивовых плантациях. Однако в настоящее время реальным способом борьбы с патогенами и вредителями является использование пестицидов – химических средств защиты растений [25, С. 248], [26, С. 96]. Современный ассортимент пестицидов достаточно обширен [27, С. 938], что дает возможность выбрать препараты, наиболее пригодные для использования на ивовых плантациях. На наш взгляд [28, С. 3], надежную защиту от стеблевых вредителей создают инсектициды с глубинным, трансламинарным и системным эффектом и препаративные формы традиционных пестицидов с пролонгированным действием [29, С. 53]:
Для того чтобы оптимизировать пестицидную нагрузку на агробиоценозы и при этом обеспечить надежную защиту интенсивных ивовых плантаций от стеблевых и иных вредителей (патогенов), необходимо разработать режимы обработки насаждений различного рода химическими средствами защиты растений [28, С. 3] с учетом ритмов нарастания годичных побегов. Необходимость изучения сезонных ритмов развития древесных растений с целью создания научно обоснованных календарных планов проведения агролесотехнических мероприятий была обоснована еще в работах Н.П. Кренке [30, С. 42]. Однако к текущему моменту сезонная динамика развития побегов ивы трехтычинковой изучена недостаточно, что затрудняет проведение таких исследований.
Цель данного исследования: выявить основные закономерности сезонной динамики нарастания побегов в модельной инбредной популяции ивы трехтычинковой.
Материал и методы
Объект исследования: модельная инбредная популяция ивы трехтычинковой, созданная путем семенной репродукции на протяжении трех поколений (в течение 2000…2017 гг.). История создания и условия культивирования данной семьи описана ранее [31, С. 22]. По состоянию на 2018 г. в состав модельной популяции входило 24 высокопродуктивных сеянца второго года жизни: tr 04… tr 24. Не исключено, что изучаемые сеянцы – F3 – имеют апомиктическое происхождение: ранее [32, С. 7] нами была обоснована гипотеза о возможности гаметофитного апомиксиса у ивы трехтычинковой в виде редуцированного партеногенеза с гаплоидностью зародышей и последующей спонтанной соматической диплоидизацией. В данном исследовании для получения материала использовались наиболее сильные сеянцы, у которых сохранялся непрерывный рост побегов без заметных повреждений. Все сеянцы формировались в один побег.
Наблюдения за линейным приростом однолетних побегов проводились на протяжении периода вегетации 2018 г. (формат даты наблюдения – день.месяц). За первый день вегетации была принята дата устойчивого перехода среднесуточных температур через 10°С (30.4), за последний день – отмирание верхушечной почки (18.7–19.8). Начиная с 07.05.2018 (при достижении побегами длины 80…200 мм), каждые четверо суток производили замеры длины побегов L (точность измерений ± 10 мм). Каждому очередному наблюдению i был присвоен порядковый номер k (1, 2, 3… k i). Все даты наблюдений (при стартовой дате 30.4), были пронумерованы по порядку: 12 (11.5), 16 (15.5), 20 (19.5)… t i.
Для дальнейшего изучения были использованы срезанные вызревшие побеги. На каждом побеге были отмечены точки, соответствующие длине побегов (L, мм) для каждой даты наблюдений t i, начиная с 07.05. Затем на каждом побеге для каждого междоузлия с порядковым номером n производили измерение его длины I, мм. Далее для каждого побега (каждого сеянца) вычисляли среднюю длину междоузлий по формуле:
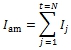 (1)
(1)
где I am – средняя арифметическая (arithmetic mean) длина междоузлий (мм) на побеге данного сеянца, I j – длина междоузлия I (мм) с порядковым номером j, N – число междоузлий на побеге.
На каждом побеге для каждой расчетной даты наблюдений tk были вычислены средние значения длины междоузлий I (t k), сформировавшихся в интервале tk–1…tk+1, по формуле:
где I (tk) – средняя арифметическая длина междоузлий (мм) для порядкового номера наблюдений k в интервале от предыдущей пронумерованной даты (tk–1) до последующей пронумерованной даты (tk+1); – сумма длин междоузлий (мм) в интервале tk–1…tk–+1; n – число междоузлий в интервале tk–1…tk–+1; временной интервал tk–1…tk–+1, для которого рассчитывалась средняя длина междоузлий, составил 9 сут.
В итоге для каждого сеянца были вычислены выровненные эмпирические ряды динамики среднесуточной длины междоузлий I (t) в период с 11.5 (t = 12) по 15.8 (t = 108). Далее на основании рядов I (t), вычисленных для каждого побега, вычисляли эмпирические ряды динамики отклонений фактических значений длины междоузлий от средней арифметической длины междоузлий (3):
![]() (3)
(3)
где dI (t) – отклонение I (t) от I am, I (t) – фактический показатель I (t k), I am – средняя арифметическая длина междоузлий; единицы измерения, мм.
На основании эмпирических рядов dI (t) вычисляли средний (агрегированный) ряд для всей выборки. Эмпирические ряды динамики dI (t) каждого побега были аппроксимированы суммами гармоник h с порядковым номером ν (4):где dI(t)calc – ожидаемое отклонение фактических значений длины междоузлий от средней арифметической длины междоузлия, ν – порядковый номер гармоники; A ν – амплитуда колебания; π – число «пи»; t – пронумерованные даты наблюдений; T – главный период колебаний, φ ν – начальная фаза колебаний в интервале 0…2 π (в дальнейшем преобразованная в sin φ), ε ν – остаточное смещение колебания по амплитуде (обусловленное ошибкой определения главного периода колебаний); период колебания каждой гармоники равен τ = T/ν.
Подробное описание изложенного алгоритма гармонического анализа дано в наших предыдущих работах [31, С. 22]. Достоверность аппроксимации эмпирических рядов каждой гармоникой оценивали с помощью F-критерия Фишера. Достоверность аппроксимации эмпирических рядов суммой гармоник определяли, сравнивая расчетные (calc) и критические (crit) значения F-критерия. Адекватность аппроксимации эмпирических рядов суммами гармоник оценивали с помощью коэффициента детерминации R2. На основании амплитудно-частотных и фазово-частотных характеристик сезонной динамики отклонений фактических значений длины междоузлий от средней арифметической длины – A (ν) и sin φ (n) – были выявлены основные биоритмы, определяющие цикличность сезонной динамики длины междоузлий.
Результаты и обсуждение
В течение периода вегетации велись наблюдения за нарастанием побегов наиболее сильных сеянцев: tr 01…tr 24. Для окончательного анализа было отобрано 9 сеянцев (tr 04, tr 05, tr 09, tr 13, tr 15, tr 17, tr 19, tr 21, tr 23), на которых длина однолетних побегов к моменту завершения роста составила 184…214 см.
Эмпирические ряды сезонной динамики отклонений фактических значений длины междоузлий от средней арифметической длины – dI (t), мм – представлены в таблице 1.
Таблица 1 – Фактическая сезонная динамика отклонений фактических значений длины междоузлий от средней арифметической длины
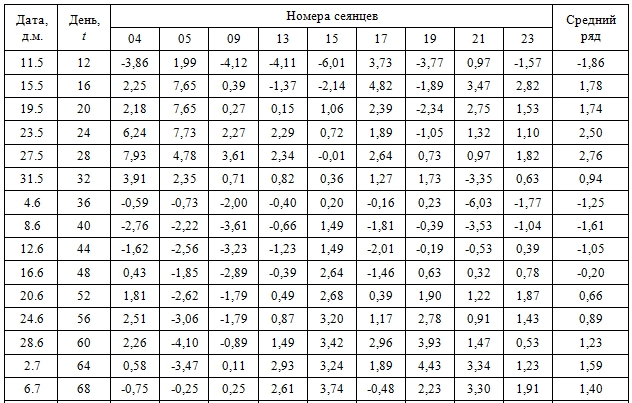
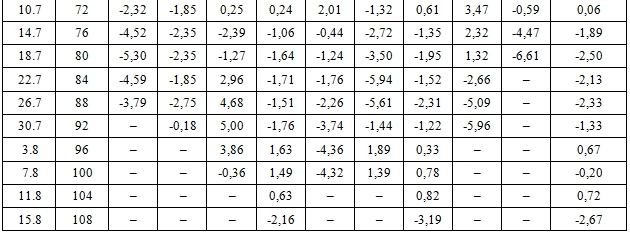
Сравнительный анализ эмпирических рядов dI (t) показал, что с середины мая до середины августа отчетливо выделяется три периода роста с максимальной длиной междоузлий: весенний (15.5…31.5), среднелетний (20.6...10.7) и позднелетний (30.7…11.8). Средний интервал между максимумами составил 36 сут. Выявленные максимумы длины междоузлий разделены двумя минимумами: раннеиюньским (4.6…8.6: исключение побег сеянца tr 05) и позднеиюльским (18.7…26.7). Интервал между минимумами составил 48 сут.
Эмпирические ряды dI (t) с высокой надежностью аппроксимируются суммами гармоник h 0,5… h 14 при Т = 144 сут. Расчетные значения критерия Фишера (Fcalc) составили Fcalc = 26,24…89,86 при Fcrit = 3,92 (P < 0,001); при этом значения коэффициентов детерминации составили R2 = 0,962…0,989.
Амплитудно-частотные характеристики рядов dI (t) приведены в таблице 2.
Таблица 2 – Амплитудно-частотные характеристики сезонной динамики отклонений фактических значений длины междоузлий от средней арифметической длины
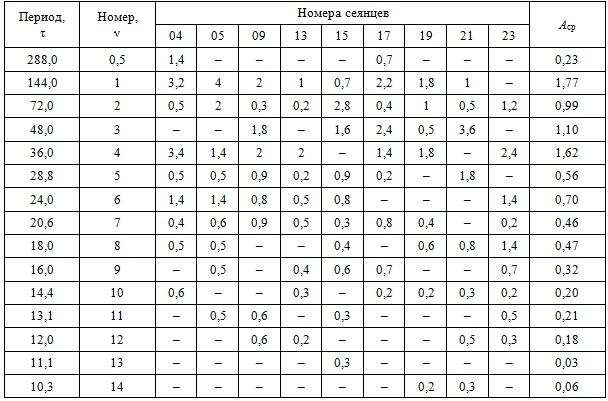
Примечание. τ – расчетные периоды колебаний гармоник, ν – номера гармоник; в ячейках указаны значения коэффициентов А ν уравнений синусоид гармоник h 0,5… h 14 для каждого побега и средние арифметические значения Аср.
Для выявления синхронности колебаний в гармониках различного порядка были использованы фазово-частотные характеристики рядов dI (t) (табл. 3).
Таблица 3 – Фазово-частотные характеристики сезонной динамики отклонений фактических значений длины междоузлий от средней арифметической длины
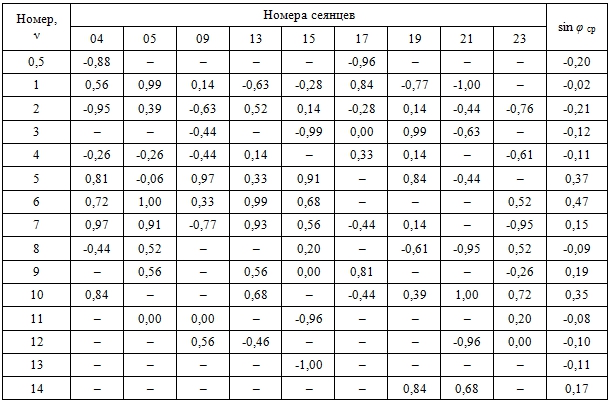
Примечание. ν – номера гармоник; в ячейках указаны значения синусов начальных фаз колебаний φ ν уравнений синусоид гармоник h 0,5… h 14 для каждого побега и средние арифметические значения sin φ ср.
Сравнительный анализ амплитудно-частотных спектров показал, что по значениям коэффициентов А в ряду h 0,5…h 14 выделяется три группы гармоник: низшие, средние и высшие. Первые гармоники (τ1 = 144 сут) определяют сезонный тренд динамики длины междоузлий. Вторая (τ2 = 72 сут) и третья (τ3 = 48 сут) гармоники, а также дробные гармоники (τ0,5 = 288 сут) оказывают на сезонный тренд модулирующее влияние. Четвертые гармоники (τ4 = 32 сут) на побегах большинства сеянцев (кроме tr 15 и tr 21) определяют пиковые значения всех трех максимумов I (весеннего 23.5, среднелетнего 28.6 и позднелетнего 7.7) и двух минимумов I (июньского 12.6 и июльского 18.7). Шестые гармоники (τ4 = 24 сут) оказывают существенное модулирующее влияние на сезонную динамику длины междоузлий на побегах большинства сеянцев (кроме tr 17, tr 19 и tr 21). Результирующие колебания высших гармоник (τ ≈ 10…21 сут) образуют квазициклические ряды с эмпирическими периодами колебаний 12…28 сут.
Сравнительный анализ фазово-частотных спектров показал, что низшие (h 0,5…h 3) и высшие (h 7…h 14) гармоники разных сеянцев не синхронизированы. Начальные фазы колебаний средних (h 4…h 6) гармоник образуют компактные множества, что указывает на их определенную синхронизацию.
Сравнительный анализ амплитудно-частотных и фазово-частотных спектров показал, что в большинстве случаев закономерности сезонной динамики длины междоузлий, общие для изучаемой модельной популяции, определяются взаимодействием высокоамплитудных четвертых и низкоамплитудных шестых гармоник.
Квазипериодический результирующий ряд dI (t), полученный путем суммирования четвертых и шестых гармоник (кроме сеянца tr 21), и фактический (actual) агрегированный ряд динамики dI (t) показаны на рисунке 1.
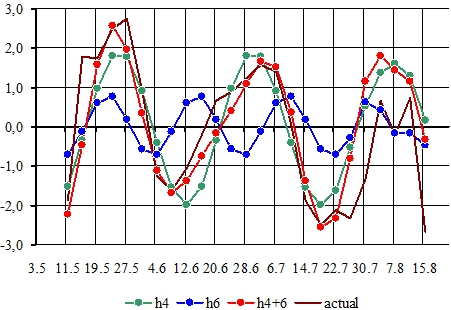
Рис. 1 – Общие компоненты сезонной динамики длины междоузлий, обусловленные взаимодействием четвертой и шестой гармоник
Примечание: по оси абсцисс даты вегетационного периода 2018 г., по оси ординат отклонения фактических значений длины междоузлий от средней арифметической длины dI (t), мм. В легенде указаны ряды гармоник h4 и h6, результирующее колебание h4+h6 и фактическая (actual) агрегированная динамика dI (t).
Побег сеянца tr 21 представляет собой исключение: максимумы сезонной динамики длины междоузлий определяются пятой гармоникой (τ ≈ 29 сут).
Выводы
Закономерности сезонной динамики длины междоузлия ивы трехтычинковой, установленные в данном исследовании, согласуются с теоретическими положениями современной биоритмологии [33, С. 104].
- Сезонная динамика длины междоузлия носит циклический характер. Эмпирические ряды сезонной динамики длины междоузлия представляют собой квазипериодические колебания. На протяжении периода вегетации в популяционной выборке выявлено три максимума, разделенных двумя минимумами.
- Сезонные тренды динамики длины междоузлий зависят от индивидуальных особенностей сеянцев и обусловлены взаимодействием низших несинхронизированных гармоник: первых (τ=144 сут), вторых (τ = 72 сут) и третьих (τ = 48 сут).
- Цикличность сезонной динамики длины междоузлия, не зависящая от индивидуальных особенностей сеянцев, в большинстве случаев определяется четвертыми гармониками (τ=36 сут). Шестые гармоники (τ = 24 сут) оказывают модулирующее влияние на цикличность сезонной динамики длины междоузлия.
- Высшие низкоамплитудные гармоники (τ=10…21 сут) в сумме образуют квазициклические ряды, конфигурация которых зависит от индивидуальных особенностей сеянцев. В популяционной выборке высшие гармоники оказывают незначительное модулирующее влияние на сезонную динамику длины междоузлия.
- При планировании мероприятий по использованию химических средств защиты плантаций ивы трехтычинковой от вредителей и патогенов рекомендуется учитывать, прежде всего, ритмы развития побегов с периодичностью колебаний порядка 36сут. Трехкратная обработка насаждений различного рода химическими средствами защиты растений позволит оптимизировать пестицидную нагрузку в насаждениях ивы трехтычинковой интенсивного типа.
| Конфликт интересов Не указан. | Conflict of Interest None declared. |
Список литературы / References
- Skvortsov A.K. Willows of Russia and adjacent countries. Taxonomical and geographical revision / A.K. Skvortsov – Joensuu: University of Joensuu, 1999. – 307 p.
- Валягина-Малютина Е.Т. Ивы европейской части России / Е.Т. Валягина-Малютина. – М.: КМК, 2004. – 217 с.
- Баркалов В.Ю. Филогенетические отношения видов Salix L. subg. Salix (Salicaceae) по данным секвенирования межгенных спейсеров хлоропластного генома и ITS ядерной рибосомальной ДНК / В.Ю. Баркалов, М.М. Козыренко // Генетика. – 2014. – Т. 50. – № 8. – С. 940–949.
- Blackburn K. A preliminary account of the chromosomes and chromosome behavior in the Salicaceae / K. Blackburn, J. Harrison // Ann. Bot. – 1924. – 38. – P. 361–378.
- Wilkinsson J. The cytology of Salix in relation to its taxonomy / J. Wilkinsson // Ann. Bot. NS. – 1944. – P. 269–283.
- Neumann A. Cytotaxonomischer Beitrag zur Gattung Salix / A. Neumann, A. Polatschek // Ann. Naturhistor. Mus. – 1972. – 76. – S. 619-633.
- Hörandl E. Die Gattung Salix in Österreich mit Berücksichtigung angrenzender Gebiete / E. Hörandl // Abh. der Zool.-Bot. Ges. – 1992. – 27. – 170S.
- Афонин А.А. Изменчивость длины пыльцевых зерен аборигенных видов ив Брянского лесного массива / А.А. Афонин // Известия высших учебных заведений. Лесной журнал. – 2006. – № 5. – С. 25–34.
- Цыганов А.Р. Биоэнергетика: энергетические возможности биомассы / А.Р. Цыганов, А.В. Клочков. – Минск: Беларус. навука, 2012. – 143с.
- Фучило Я.Д. Створення та вирощування енергетичних плантацій верб i тополь: Науково-методичнi рекомендації / Я.Д. Фучило, М.В. Сбитна, О.Я. Фучило, В.М. Лiтвiн. – К.: Логос, 2009. – 80 с.
- Karp A. Genetic improvement of willow for bioenergy and biofuels / A. Karp, S.J. Hanley, S.O. Trybush [et al.] // Journal of integrative plant biology. – 2011. – 53(2). – P. 151–165. – DOI: https://doi:10.1111/j.1744-7909.2010.01015.x
- Гомилевский В.И. Культура ивы и корзиночное производство / В.И. Гомилевский. – СПб.: Книгоизд-во П.П. Сойкина, 1907. – 41 с.
- Керн Э.Э. Ива, ее значение, разведение и употребление / Э.Э. Керн. – 4-е изд., вновь перераб. – Петроград: Тип. Мин-ва Путей Сообщения (Тов-ва И.Н. Кушнерев и К°), 1915. – 134с.
- Kuzovkina Y.A. Checklist for Cultivars of Salix (Willow) / Y.A. Kuzovkina // International Salix Cultivar Registration Authority. – 2015. – 50. – P. 163.
- Mikó P. Biomass production of energy willow under unfavourable field conditions / P. Mikó, G.P. Kovács, L. Alexa [et al.] // Applied Ecology and Environmental Research. – 2014. – 12(1). – P. 1–12.
- Фучило Я.Д. Рост и продуктивность некоторых сортов энергетической ивы в зависимости от степени увлажненности почвы / Я.Д. Фучило, М.В. Сбытна, Б.В. Зелинский // Plant Varieties Studying and Protection. – 2018. – 14(3). – С. 323-327.
- Афонин А.А. Ивы Брянского лесного массива: проблема повышения продуктивности и устойчивости насаждений и пути ее решения / А.А. Афонин. – Брянский гос. ун-т, 2005. – 172 с. – Деп. в ВИНИТИ 15.09.05, № 1227–В2005. – Библиогр. указатель «Депонир. науч. работы». – № 11. – 2005.
- Niemi L. Spatial variation in resistance and virulence in the host-pathogen system Salix triandra–Melampsora amygdalinae / Niemi, A. Wennström, J. Hjältén [et al.] //Journal of Ecology. – 2006. – 94. – P. 915–921. – DOI: https://doi:10.1111/j.1365-2745.2006.01157.x
- Rönnberg-Wästljung A.C. Resistance to Melampsora larici-epitea leaf rust in Salix: analyses of quantitative trait loci / A.C. Rönnberg-Wästljung, B. Samils, V. Tsarouhas, U. Gullberg // Journal of Applied genetics. – 2008. – 49. – P. 321–331.
- Sage R.B. The distribution of Phratora vulgatissima (Coleoptera: Chrysomelidae) on cultivated willows in Britain and Ireland / R.B. Sage, K. Tucker // Forest Pathology. – 1998. – 28. – P. 289‑
- Torp M. Performance of an Herbivorous Leaf Beetle (Phratora vulgatissima) on Salix F2 Hybrids: the Importance of Phenolics / M. Torp, A. Lehrman, J.A. Stenberg [et al.] // Journal of Chemical Ecology. – 2013. – 39(4). – P. 516–524. – DOI: http://dx.doi.org/10.1007/s10886-013-0266-3
- Czesak M.E. Genetic architecture of resistance to aphids and mites in a willow hybrid system / M.E. Czesak, M.J. Knee, R.G. Gale and others // Heredity. – 2004. – 93. – P. 619–626.
- Ollerstam O. Salicylic acid mediates resistance in the willow Salix viminalis against the gall midge Dasineura marginemtorquens / O. Ollerstam, S. Larson // Journal of Chemical Ecology. – 2003. – 29. – P. 163–174.
- Афанасенко О.С. Генетическая защита растений: проблемы и перспективы / О.С. Афанасенко // Защита и карантин растений. – 2016. – 1. – С.13–16.
- Ганиев М.М. Химические средства защиты растений / М.М. Ганиев, В.Д. Недорезков. – М.: КолосС, 2006. – 248 с.
- Попова Л.М. Химические средства защиты растений: Учебное пособие / Л.М. Попова. – СПб.: СПбГТУРП, 2009. – 96 с.
- Государственный каталог пестицидов и агрохимикатов, разрешенных к применению на территории Российской Федерации. – Ч.1. – Пестициды. – Издание официальное. – М.: Минсельхоз РФ, 2017. – 938 с.
- Афонин А.А. Генетическая безопасность агробиоценозов / А.А. Афонин, Л.И. Булавинцева // Биология в школе. – 2011. – №5. – С. 3–10.
- Литвишко В.С. Экологизация средств защиты растений / В.С. Литвишко // Естественные и технические науки. – 2018. – № 2 (116). – С. 53-54.
- Кренке Н.П. Теория циклического старения и омоложения растений и практическое её применение / Н.П. Кренке. – М.: Огиз–Сельхозгиз, 1940. – 135с.
- Афонин А.А. Структурный анализ ритмов развития однолетних побегов ивы трехтычинковой / А.А. Афонин // Бюлл. науки и практики. – – Т. 5. – №1. – С. 22–32.– DOI:http://doi.org/10.5281/zenodo.2539541
- Афонин А.А. Изменчивость морфологии листовых пластинок при семенной репродукции ивы трехтычинковой / А.А. Афонин // Междунар. науч.-иссл. журн. – 2018. – №9(75). – Ч.2. – С. 7–13. – DOI: https://doi.org/10.23670/IRJ.2018.75.9.025
- Ежов С.Н. Основные концепции биоритмологии / С.Н. Ежов // Вестн. ТГЭУ. – 2008. – № 2. – С. 104–121.
Список литературы на английском языке / References in English
- Skvortsov A.K. Willows of Russia and adjacent countries. Taxonomical and geographical revision / A.K. Skvortsov – Joensuu: University of Joensuu, 1999. – 307 p.
- Valyagina-Malyutina E.T. Ivy evropejskoj chasti Rossii [Willows of the European part of Russia] / E.T. Valyagina-Malyutina. – M.: KMK, 2004. – 217 p. [in Russian]
- Barkalov V.Yu. Filogeneticheskie otnosheniya vidov Salix subg. Salix (Salicaceae) po dannym sekvenirovaniya mezhgennyh spejserov hloroplastnogo genoma i ITS yadernoj ribosomal'noj DNK [Phylogenetic relations of Salix L. subg. Salix species (Salicaceae) according to sequencing data of intergenic spacers of chloroplasic genomes and ITS rDNA] / V.Yu. Barkalov, M.M. Kozyrenko // Genetika [Russian Journal of Genetics]. – 2014. – 50(8). – P. 940–949. [in Russian]
- Blackburn K. A preliminary account of the chromosomes and chromosome behavior in the Salicaceae / K. Blackburn, J. Harrison // Ann. Bot. – 1924. – 38. – P. 361–378.
- Wilkinsson J. The cytology of Salix in relation to its taxonomy / J. Wilkinsson // Ann. Bot. NS. – 1944. – P. 269–283.
- Neumann A. Cytotaxonomischer Beitrag zur Gattung Salix / A. Neumann, A. Polatschek // Ann. Naturhistor. Mus. – 1972. – 76. – P. 619-633.
- Hörandl E. Die Gattung Salix in Österreich mit Berücksichtigung angrenzender Gebiete / E. Hörandl // Abh. der Zool.-Bot. Ges. – 1992. – 27. – 170 P.
- Afonin A.A. Izmenchivost' dliny pyl'cevyh zeren aborigennyh vidov iv Bryanskogo lesnogo massiva [Variability of Pollen-grains Length for Native Species of Willows in Bryansk Woodland] / A.A. Afonin // Izvestiya vysshih uchebnyh zavedenij. Lesnoj zhurnal [Bulletin of higher educational institutions. Forestry Journal]. – 2006. – 5. – P. 25–34. [in Russian]
- Cyganov A.R. Bioenergetika: energeticheskie vozmozhnosti biomassy [Bioenergy: energy potential of biomass] / A.R. Cyganov, A.V. Klochkov. – Minsk: Belarus. navuka, 2012. – 143 p. [in Russian]
- Fuchilo Ya.D. Stvorennya ta viroshchuvannya energetichnih plantacіj verb i topol': Naukovo-metodichni rekomendacії [The creation and cultivation of energy plantations of willows and poplars. Scientific-methodical recommendations] / Ya.D. Fuchilo, M.V. Sbitna, O.Ya. Fuchilo, V.M. Litvin. – K.: Logos, 2009. – 80 p. [in Ukraine]
- Karp A. Genetic improvement of willow for bioenergy and biofuels / A. Karp, S.J. Hanley, S.O. Trybush [et al.] // Journal of integrative plant biology. – 2011. – 53(2). – P. 151–165. – DOI: https://doi:10.1111/j.1744-7909.2010.01015.x
- Gomilevskij V.I. Kul'tura ivy i korzinochnoe proizvodstvo [The culture of willow and basket manufacturing] / V.I. Gomilevskij. – SPb.: Knigo Publishing house P.P. Sojkina, 1907. – 41 p. [in Russian]
- Kern E.E. Iva, ee znachenie, razvedenie i upotreblenie [Willow, its value, and breeding and consumption] / E.E. Kern. – 4-e edition., vnov' pererab. – Petrograd: Tip. Min-va Putej Soobshcheniya (Tov-va I.N. Kushnerev i K°), 1915. – 134 p. [in Russian]
- Kuzovkina Y.A. Checklist for Cultivars of Salix (Willow) / Y.A. Kuzovkina // International Salix Cultivar Registration Authority. – 2015. – 50. – P. 163.
- Mikó P. Biomass production of energy willow under unfavourable field conditions / P. Mikó, G.P. Kovács, L. Alexa [et al.] // Applied Ecology and Environmental Research. – 2014. – 12(1). – P. 1–12.
- Fuchilo Ya.D. Rost i produktivnost' nekotoryh sortov energeticheskoj ivy v zavisimosti ot stepeni uvlazhnennosti pochvy [Growth and productivity of some cultivars of energy willow, depending on the degree of soil moisture] / Ya.D. Fuchilo, M.V. Sbytna, B.V. Zelinskij // Plant Varieties Studying and Protection. – 2018. – 14(3). – P. 323-327. [in Ukraine]
- Afonin A.A. Ivy Bryanskogo lesnogo massiva: problema povysheniya produktivnosti i ustojchivosti nasazhdenij i puti ee resheniya [The willows of the Bryansk Woodland: the problem of increasing the productivity and stability of plantations and ways to solve it] / A.A. Afonin. – Bryanskij gos. un-t, 2005. – 172 p. – Dep. v VINITI 15.09.05, № 1227–V2005. – Bibliogr. ukazatel' «Deponir. nauch. raboty». – 11. – 2005. [in Russian]
- Niemi L. Spatial variation in resistance and virulence in the host-pathogen system Salix triandra–Melampsora amygdalinae / Niemi, A. Wennström, J. Hjältén [et al.] //Journal of Ecology. – 2006. – 94. – P. 915–921. – DOI: https://doi:10.1111/j.1365-2745.2006.01157.x
- Rönnberg-Wästljung A.C. Resistance to Melampsora larici-epitea leaf rust in Salix: analyses of quantitative trait loci / A.C. Rönnberg-Wästljung, B. Samils, V. Tsarouhas, U. Gullberg // Journal of Applied genetics. – 2008. – 49. – P. 321–331.
- Sage R.B. The distribution of Phratora vulgatissima (Coleoptera: Chrysomelidae) on cultivated willows in Britain and Ireland / R.B. Sage, K. Tucker // Forest Pathology. – 1998. – 28. – P. 289‑
- Torp M. Performance of an Herbivorous Leaf Beetle (Phratora vulgatissima) on Salix F2 Hybrids: the Importance of Phenolics / M. Torp, A. Lehrman, J.A. Stenberg [et al.] // Journal of Chemical Ecology. – 2013. – 39(4). – P. 516–524. – DOI: http://dx.doi.org/10.1007/s10886-013-0266-3
- Czesak M.E. Genetic architecture of resistance to aphids and mites in a willow hybrid system / M.E. Czesak, M.J. Knee, R.G. Gale [et al.] // Heredity. – 2004. – 93. – P. 619–626.
- Ollerstam O. Salicylic acid mediates resistance in the willow Salix viminalis against the gall midge Dasineura marginemtorquens / O. Ollerstam, S. Larson // Journal of Chemical Ecology. – 2003. – 29. – P. 163–174.
- Afanasenko O.S. Geneticheskaya zashchita rastenij: problemy i perspektivy [Genetic protection of plants: problems and prospects] / O.S. Afanasenko // Zashchita i karantin rastenij [Plant protection and quarantine]. – 2016. – 1. – 13–16. [in Russian]
- Ganiev M.M. Himicheskie sredstva zashchity rastenij [Chemical plant protection products] / M.M. Ganiev, V.D. Nedorezkov. – M.: KolosS, 2006. – 248 p. [in Russian]
- Popova L.M. Himicheskie sredstva zashchity rastenij: Uchebnoe posobie [Chemical plant protection products: Study guide] / L.M. Popova. – SPb.: SPbGTURP, 2009. – 96 p. [in Russian]
- Gosudarstvennyj katalog pesticidov i agrohimikatov, razreshennyh k primeneniyu na territorii Rossijskoj Federacii – Ch.1. – Pesticidy. – Izdanie oficial'noe [State catalogue of pesticides and agrochemicals authorized for use on the territory of the Russian Federation. – Part 1. Pesticide. – Official publication]. – M.: Minsel'hoz RF, 2017. – 938 p. [in Russian]
- Afonin A.A. Geneticheskaya bezopasnost' agrobiocenozov [Genetic safety of agrobiocenoses] / A.A. Afonin, L.I. Bulavintseva // Biologiya v shkole [Biology at school]. – 2011. – 5. – 3–10. [in Russian]
- Litvishko V.S. Ekologizaciya sredstv zashchity rastenij [Ecologization of plant protection products] / V.S. Litvishko // Estestvennye i tekhnicheskie nauki [Natural and technical Sciences]. – 2018. – 2 (116). – 53-54. [in Russian]
- Krenke N.P. Teoriya ciklicheskogo stareniya i omolozheniya rastenij i prakticheskoe eyo primenenie [The theory of cyclic aging and rejuvenation of plants and its practical application] / N.P. Krenke. – M.: Ogiz–Sel'hozgiz, 1940. – 135 p. [in Russian]
- Afonin A.A. Strukturnyj analiz ritmov razvitiya odnoletnih pobegov ivy trekhtychinkovoj [Structural analysis of the rhythms of development of annual shoots of almond willow] / A.A. Afonin // Byull. nauki i praktiki [Bulletin of Science and Practice]. – 2019. – 5(1). – P. 22–32. – DOI: http://doi.org/10.5281/zenodo.2539541 [in Russian]
- Afonin A.A. Izmenchivost' morfologii listovyh plastinok pri semennoj reprodukcii ivy trekhtychinkovoj [Variability of lamina morphology in seeds reproduction of almond-leaved willow] / A.A. Afonin // Mezhdunar. nauch.-issl. zhurn. [International research journal] – 2018. – 9(75). – Ch.2. – P. 7–13. – DOI: https://doi.org/10.23670/IRJ.2018.75.9.025 [in Russian]
- Ezhov S.N. Osnovnye koncepcii bioritmologii [Basic concepts of biorhythmology] / S.N. Ezhov // Vestn. TGEU [Vestnik Pacific state university of economics]. – 2008. – 2. – P. 104–121. [in Russian]