ФИЛОГЕНЕТИЧЕСКОЕ ЗНАЧЕНИЕ МИКРОСТРУКТУРНЫХ ПРИЗНАКОВ ПЕРА
ФИЛОГЕНЕТИЧЕСКОЕ ЗНАЧЕНИЕ МИКРОСТРУКТУРНЫХ ПРИЗНАКОВ ПЕРА
Научная статья
Силаева О.Л.1, *, Вараксин А.Н.2
1 Институт проблем экологии и эволюции им. А.Н. Северцова (ИПЭЭ РАН), Москва, Россия;
2 Научно-исследовательский центр распознавания образов, Москва, Россия
* Корреспондирующий автор (silaeva.o[at]gmail.com)
АннотацияС помощью однофакторного дисперсионного анализа показано, что микроструктурные особенности количественных микро-морфологических компонентов пера могут свидетельствовать о филогенетическом родстве таксонов и соответственно могут использоваться как таксономические признаки. Сегментация проводилась по отрядам Гусеобразных; Курообразных; Ржанкообразных; Голубеобразных; Дятлообразных; Воробьинообразных, а также по семействам внутри отряда Ржанкообразных. Отряд Дятлообразных по данным сегментации на основании микроструктурных перьевых признаков показал родственную связь с отрядом Воробьинообразных.
Ключевые слова: морфология пера, микроструктура пера, идентификация вида птицы по одиночному перу.
PHILOGENETIC VALUE OF THE MICROSTRUCTURAL FEATURES OF A FEATHER
Research article
Silaeva O.L.1, *, Varaksin A.N.2
1 A.N. Severtsov Institute of Ecology and Evolution, Moscow, Russia;
2 Research Centre for Pattern Recognition, Moscow, Russia
* Corresponding author (silaeva.o[at]gmail.com)
AbstractUsing single-factor analysis of variance, it was shown that the micro-structural features of the quantitative micro-morphological components of a feather can indicate phylo-genetic relationship of taxonomic units and, accordingly, can be used as taxonomic characteristics. Segmentation was carried out by such orders as the Anseriformes; Galliformes; Charadriiformes; Columbiformes; Piciformes; Passeriformes, as well as families within the order of Charadriiformes. The order of Piciformes showed a kinship with the order of Passeriforms according to the segmentation data based on the micro-structural feather characteristics.
Keywords: feather morphology, feather micro-structure, bird species identification from a single feather.
ВведениеОтдельное перо птицы представляет собой системную единицу всего оперения. При этом оно в свою очередь является сложным системным комплексом связанных между собой, коррелированных и некоррелированных в линейно-метрических соотношениях микро- и макроструктурных элементов. Нам пока очень мало известно о внутреннем системном характере строения, соотношениях этих двух систем, а также о зависимостях между отдельными элементами внутри одиночного пера птицы.
Работы по выявлению микроструктурных отличий перьевых структур, имеющих таксономическое значение, ведутся не первый год. Классические исследования зарубежных авторов [20], [23], [25] определили место морфологии пера (птилологии), как научного направления в области таксономии наряду с морфологией других органов животных и остеологией. Практическая необходимость выявления морфологических микроструктурных отличий перьевых структур в связи с идентификационными работами в области авиационной орнитологии, в частности, привела к созданию мультимедийной сводки по идентификации останков птиц [26], [27].
В 1960–1970-х гг. исследования перьевого материала начали проводить учёные академической и отраслевой науки. Исследования морфологии пера, его адаптивных, функциональных и таксономических структурных особенностей было возобновлено в ИПЭЭ РАН [14]. Наряду с изучением качественных характеристик перьевых структур был начат анализ количественных признаков.
Впервые количественные характеристики признаков архитектоники пера были исследованы на основе статистического анализа и результаты исследований показали возможность использования признаков архитектоники пера в качестве таксономических [21].
Цель данной работы – показать насколько микроструктурные элементы пухового луча могут использоваться для определения филогенетических связей между высшими таксонами (отряды, подотряды и семейства), а также служить диагностическими признаками при идентификации этих классов. Одной из основных задач было выявление наиболее информативных и некоррелированных признаков пухового луча и объединение их в систему.
Материал
Исследовались пуховые структуры полупуховых покровных, преимущественно межлопаточных перьев (tectrices interscapularis) (см. рисунок 1).
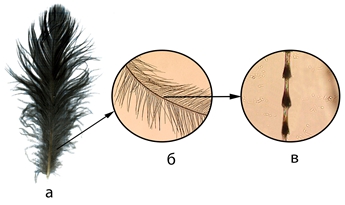
Рис. 1 – Межлопаточное перо сороки Pica pica – а; бородка – б; луч – в
Выборка, представленная в виде таблицы из 4060 строк, каждая из которых содержит значения шести признаков, включает следующие отряды с семействами, родами и видами.
- Гусеобразные − одно семейство, девять родов, два вида;
- Курообразные − двумя семействами, девятью родами, девятью видами;
- Ржанкообразные – шестью семействами, 21 родом, 35 видами;
- Голубеобразные − одним семейством, двумя родами, пятью видами;
- Дятлообразные − одним семейством, двумя родами, тремя видами;
- Воробьинообразные представлены семью семействами, 16 родами, 23 видами.
Методы
В работе применяли следующие методы.
- Светооптическая микроскопия: микроскоп Leica DMR 2700 (Leica, Германия), оснащенный фотокамерой JVC 3 CCD C-MOUNT, Измерения проводили с использованием программы LeicaQWin. Обработку изображений проводили в программе Adobe Photoshop CC 2014.
- Препарирование и микро-морфометрия.
- Создание выборки.
- Однофакторный дисперсионный анализ.
Метод микро-морфометрии
Для создания выборки проводились измерения пуховых структур (рис.1, табл. 1) по оригинальному авторскому методу. Из медиальных участков двух–трех бородок выбирали лучи с целыми заостренными, а значит неповреждёнными вершинами. Делали по 10 промеров каждого из шести указанных в таблице 2 признаков. На каждую особь таким образом приходилось по 60 измерений, а вид характеризовался, по крайней мере, 180 промерами. Измерения проводили при увеличении 400× (см. рисунок 2).
Таблица 1 – Морфометрические признаки элементов типичного пухового луча
| № признака | Признак (структура), ед. измерения | Сокращенное наименование признака |
| 1 (P1) | Среднее количество узлов на 1 мм длины луча (плотность узлов), шт. | ρузл |
| 2 (P2) | Длина междоузлия, мкм | lмежд |
| 3 (P3) | Ширина междоузлия, мкм | wмежд |
| 4 (P4) | Длина узла, мкм | lузл |
| 5 (P5) | Ширина узла, мкм | wузл |
| 6 (P6) | Длина типичного пухового луча, мм | lлуча |

Рис. 2 – Морфометрия пуховых лучей на примере лучей межлопаточного пера краснозобика
Примечание: Условные обозначения см. в таблице 1
Результаты и обсуждениеФормирование выборки для проведения статистического анализа
Эффективное решение задачи по выявлению филогенетических связей между таксонами на основе микроструктурных признаков пера напрямую связано с выбором информативных и некоррелированных между собой признаков. Репрезентативная выборка, описывающая все разнообразие особей через признаки, т.е. через микроструктурные особенности перьевых структур, является важнейшим элементом статистического анализа.
В качестве диагностически значимых перьевых микроструктур в конце XX столетия были предложены два признака – длина пухового луча базальной части пера и плотность узлов на этом луче (см. рисунок 2). Их с разной долей успеха применяли для идентификации высших таксонов [16], [26]. Пуховые структуры для измерений брали от одного пера, принадлежащего одной особи вида. При этом изученные перья принадлежали разным птерилиям: грудинным, брюшным, затылочным, кроющим хвоста и в некоторых случаях даже мантии. Однако отмечалось, что микроструктурный анализ нужно проводить по перьям одной части тела, то есть признавался факт наличия вариативности перьевых структур разных птерилий [17], [18].
Двух признаков было недостаточно для определения места каждого таксона в системе, а другими словами – для выявления филогенетических связей между таксонами. Мы дополнили имеющиеся признаки четырьмя новыми и получили шесть количественных признаков (см. таблицу 1, рисунок 2). Часть из них упоминались в литературе, но использовались редко и бессистемно, статистический анализ на их основе не проводился. Мы сформировали систему признаков, составившую единый морфологический комплекс одного пухового луча; этот комплекс по нашему мнению отражает специфику таксона (см. таблицу 1, рисунок 2).
При этом за бортом остались некоторые важные признаки пуховой части пера, данные по которых пока не удалось преобразовать в количественные. Это в частности, узловые зубцы; при наличии критерия количественной оценки этих элементов луча их вероятно можно было бы использовать как признак.
Статистический анализ
Информативность признаков. В качестве статистического анализа был применён однофакторный дисперсионный анализ. Прежде, чем приступить к собственно анализу, требуется определить информативность признаков, что является необходимым условием получения количественной оценки признаков при разбиении выборки на заданное число сегментов (отряд, семейство, род, вид).
Под информативностью понимается способность признака принимать одни количественные значения на элементах одного класса и совершенно другие на элементах другого класса [5]; чем выше информативность, тем выше способность признака отличить элементы одного класса от элементов другого класса и наоборот. С помощью однофакторного дисперсионного анализа мы оцениваем степень информативности признаков и возможность их дальнейшего использования в качестве диагностических и для определения родственных связей между таксонами.
Информативность h признака ![]() рассчитывается по формуле:
рассчитывается по формуле:
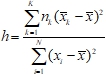 (1)
(1)
![]() матожидание k-го класса и всей выборки соответственно, nk – количество элементов к-го класса распознавания, а N – размерность исходного признакового пространства. Информативность h является безразмерной величиной, область ее определения находится в диапазоне от 0 до 1. При этом информативность h=1 означает, что значения класса k1 существенно отличаются от класса kn.
матожидание k-го класса и всей выборки соответственно, nk – количество элементов к-го класса распознавания, а N – размерность исходного признакового пространства. Информативность h является безразмерной величиной, область ее определения находится в диапазоне от 0 до 1. При этом информативность h=1 означает, что значения класса k1 существенно отличаются от класса kn.
Корреляция признаков. Условие, предъявляемое к признакам – отсутствие между ними линейных связей. Расчет (i, j) элемента симметричной матрицы R осуществляется по формуле:
C = cov(X) – матрица ковариаций, X –выборка, столбцами которой являются признаки, строчками – данные измерений. По причине большой размерности мы не можем привести здесь данные выборки.
Результатом расчета является симметричная матрица R сравнений пар, на диагонали которой размещены все единицы (см. таблицу 2).
Сегментация выборки. В идеале количественные признаки, используемые для сегментации, должны быть некоррелированными и информативными. Оба эти фактора – залог успешного разбиения выборки на обособленные сегменты. Наличие информативных некоррелированных признаков позволяет визуализировать сегменты данных в пространстве двух и более компонент и наглядно оценить качество сегментации выборки.
При различной сегментации выборки (группирование по разным таксонам) меняется не только количество классов, но и сила линейной связи между признаками и, конечно же, информативность. Обособленные в выбранном пространстве сегменты позволяют сформировать эталоны, по которым будут определяться родственные связи между таксонами, а также решаться задача диагностики таксона.
Сегментация выборки по отрядам. Определим коэффициенты корреляции между данными шести микроструктурных признаков, вошедшими в выборке. Довольно значительная линейная связь обнаруживается между признаками P1 и P2 (см. таблицу 2). Это повлияет на информативность и соответственно на визуализацию данных в пространстве этих двух признаков.
Таблица 2 – Корреляция между признаками при сегментации выборки по отрядам
| P1 | P2 | P3 | P4 | P5 | P6 | |
| P1 | 1 | |||||
| P2 | -0.8397 | 1 | ||||
| P3 | -0.4119 | 0.4912 | 1 | |||
| P4 | 0.4421 | -0.5775 | -0.1744 | 1 | ||
| P5 | -0.1521 | 0.1348 | 0.0637 | -0.0426 | 1 | |
| P6 | -0.3416 | 0.2777 | 0.0281 | -0.1526 | 0.6789 | 1 |
Таблица 3 – Информативность признаков при сегментации выборки по отрядам
| Признак | P1 | P2 | P3 | P4 | P5 | P6 |
| Информативность | 0.6641 | 0.8145 | 0.3729 | 0.4124 | 0.6078 | 0.7564 |
При этом максимально информативными оказались признаки P2 и P6 (см. таблицу 3), с помощью которых и удалось разбить выборку на отряды. Кроме того, эти признаки слабо коррелированы (см. таблицу 2). В пространстве двух информативных признаков P2 и P6 хорошо группируются все шесть отрядов (см. рисунок 3).
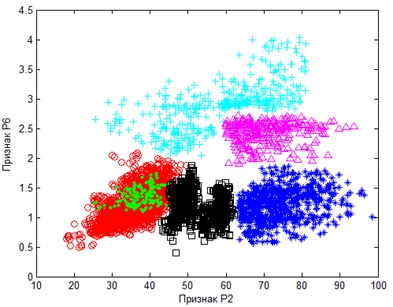
Рис. 3 – Визуализация отрядов в пространстве некоррелированных информативных признаков P2 и P6.
Условные обозначения: Гусеобразные – *; Курообразные – +; Ржанкообразные – ; Голубеобразные – ∆; Дятлообразные – зелёные точки; Воробьинообразные – ○
Все отряды довольно компактны, кроме отряда Курообразных, данные которого рассредоточены и пересекаются с кластером Голубеобразных.
Отряд Ржанкообразных находится между отрядом Воробьинообразных и отрядом Гусеобразных. В его кластере видны контуры двух кластеров подотрядов Чайковых и Ржанковых, и это чётко проявится при сегментации внутри таксона Ржанкообразных (см. рисунок 4).
Отряд Дятлообразных оказался внутри кластера отряда Воробьинообразных, показав родственную связь с ним. Получается, что по данным сегментации на основании микроструктурных перьевых признаков отряд Дятлообразных не является самостоятельным. Дискуссии по поводу филогении Дятлообразных ведутся уже в течение многих лет; в основном принимались во внимание остеологические, морфологические и другие данные. Перьевые структуры впервые были рассмотрены в качестве таксономических несколько десятилетий назад и показали свою значимость в качестве таксономических критериев. Было выявлено морфологическое сходство оперения представителей Воробьинообразных и Дятлообразных [18], [22] на уровне микроструктур: ворсинок (наличие и форма), узловых структур, пигментации и т.п. Ворсинки встречаются в отряде Воробьинообразных и в четырёх семействах Дятлообразных [19]. Сходство микроструктур Воробьинообразных и Дятлообразных продемонстрировано и в наших работах [12]. Результаты этих исследований значительно ослабили гипотезу монофилии Дятлообразных.
Сегментация отряда Ржанкообразных по подотрядам и семействам. Отряд Ржанкообразных обладает сложной филогенетической структурой, он обширен и разнообразен, отряд, состоит из трех довольно самостоятельных подотрядов: Куликов, Чайковых и Чистиковых. Для определения линейных связей между всеми шестью признаками построим по отработанной методике матрицу корреляции для отряда (см. таблицу 4).
Таблица 4 – Корреляция между признаками для отряда Ржанкообразных
| P1 | P2 | P3 | P4 | P5 | P6 | |
| P1 | 1 | |||||
| P2 | -0.6753 | 1 | ||||
| P3 | 0.1583 | -0.2808 | 1 | |||
| P4 | -0.0349 | -0.2403 | 0.1139 | 1 | ||
| P5 | 0.2208 | -0.3161 | 0.5347 | 0.1469 | 1 | |
| P6 | -0.1765 | -0.0888 | 0.2979 | 0.2169 | 0.1816 | 1 |
Коэффициенты корреляции в целом довольно низкие. Так, между признаками P1 и P2 корреляция составила 0,67, а между признаками P5 и P3 – 0,53 (см. таблицу 4). Это позволяет утверждать, что отрицательное влияние линейных связей, выявленных между признаками, будет минимально влиять на результаты разбиения в пространстве изученных признаков. Рассчитываем информативность признаков для шести семейств: Ржанковых, Бекасовых, Тиркушковые, Чайковых, Поморниковых и Чистиковых (см. таблицу 5).
Таблица 5 – Информативность признаков отряда Ржанкообразных при сегментации по семействам
| Признак | P1 | P2 | P3 | P4 | P5 | P6 |
| Информативность | 0.2192 | 0.8632 | 0.1213 | 0.1823 | 0.1117 | 0.1886 |
Анализ результатов оценки информативности признаков указывает на то, что семейства отряда Ржанкообразных наилучшим образом различаются по признаку P2 и P1 (см. рисунок 4).
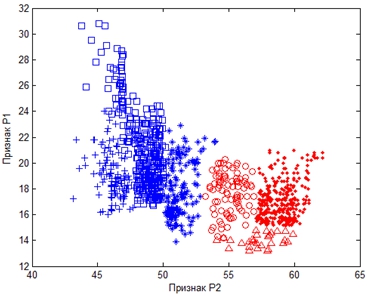
Рис. 4 – Визуализация семейств отряда Ржанкообразных в пространстве информативных признаков P1 и P2
Условные обозначения: Ржанковые – *; Бекасовые – ; Тиркушковые – +; Поморниковые – ○; Чайковые – ■; Чистиковые – ∆
Все шесть семейств достаточно компактны, за исключением Бекасовых, которые значительно разбросаны по признаку P1. В пространстве двух наиболее информативных признаков P1 и P2 информативность признака P1 составила 0,22 (см. таблицу 5). Тем не менее, на рисунке 4 отобразились кластеры шести семейств отряда Ржанкообразных; удается также увидеть обособленность подотрядов Куликов, Чайковых и Чистиковых. Семейство Поморниковых отделилось от Чайковых, хотя кластеры не выглядят четко очерченными. Разнородность отряда и обособленность подотрядов находят свое отражение и в структурных особенностях пера (см. рисунок 4).
Подотряд Чистиковых, представленный в выборке всего одним родом Cyclorrhynchus, группируется в одном кластере рядом с семейством Чайковых. Возможно, это обусловлено малочисленностью выборки по семейству Чистиковых. При более представительной выборке информативность признака Р1 могла бы быть выше и кластер по этому признаку сместился бы вниз. Тогда и отношение кластера Чистиковых к таковому Чайковых было бы довольно условным. Результаты сегментации, таким образом, позволяют утверждать, что в определении отряда, подотрядов и семейств не должно представлять существенных сложностей.
Проблемы микроморфометрии и создания выборок. Пока мы не смогли провести сегментацию выборки по родам и видам, что обусловлено многими причинами, на которых кратко остановимся ниже.
Изменчивость объектов измерения. Размеры и форма элементов изменяются в зависимости от их положения на луче. Длина луча и другие его параметры изменяются в зависимости от положения на опахальце бородки. Перья на выбранной нами межлопаточной птерилии имеют всё-таки разный размер, который влияет и на размер внутренних элементов пера, в частности, бородок и лучей соответственно [12]. Во избежание погрешностей в промерах, мы измеряли узлы и междоузлия из медиальной части луча. Лучи также выбирали из медиальной части бородки. В результате нам удалось до какой-то степени сгладить погрешности в измерениях, но не избежать их полностью.
Микроструктурные элементы некоторых видов, в частности, сильно редуцированные узлы и едва заметные выступы вместо узловых зубцов, оказались недоступными для измерения в программе LeicaQWin. Поэтому пришлось отказаться от измерения структур люрика, чистика, толстоклювой кайры, большой конюги и вальдшнепа. В результате эти виды не были включены в выборку. Однако слабую выраженность микроструктур этих видов можно использовать как признак для системы диагностических признаков. Из представителей Чистиковых удалось измерить только белобрюшку, что повлияло на полноту выборки по этому подотряду.
Малые выборки. Мы максимально повысили репрезентативность выборки путём увеличения числа промеров для одной особи. Однако в силу трудоемкости работы по измерениям нам не удалось довести выборку до средних размеров за счет увеличения числа видов. Отметим также, что оперение не всех видов было нам доступно. В отсутствии представительной статистики повышаются требования к математическим методам. Становится актуальным выбор и обоснование метрического пространства, в котором хорошо различаются рода и виды.
Погрешности в промерах. В процессе работы над созданием выборки нам стало ясно, что чем ниже таксон, тем выше требования к точности измерения микроструктурных элементов. В связи с этим более остро встает задача минимизации субъективизма операторов, ведущих выборку. Несмотря на то, что промеры выполнялись минимальным количеством операторов, измерения для одной особи не компактны.
Заключение
Несмотря на изменчивость морфологических качественных и количественных показателей у представителей разных таксономических групп оказалось возможным выявление признаков, показавших как различия между таксономическими группами, так и общие черты, которые их объединяют. Проведенная дифференциация на уровне высших таксонов свидетельствует о том, что сходство или несходство микроструктурных элементов и дополняющих их характеристик можно использовать в филогенетических исследованиях наряду с остеологическими, биохимическими и другими морфологическими данными. Дифференциация пока проведена лишь на уровне отрядов и семейств. Однако, с помощью статистического анализа на основе оценки информативности признаков удалось определить не только видовые, но и внутривидовые связи [13]. К сожалению, птилологический метод неклассической систематики, основанный на морфометрии, пока недостаточно разработан, и по этой причине почти не привлекается для исследований в области таксономии и филогенетики.
| Финансирование Работа по изучению микроструктуры проводилась с использованием оборудования Центра коллективного пользования «Инструментальные методы в экологии» при ИПЭЭ РАН. | Funding The study of microstructure was conducted using Joint Usage Center «Instrumental methods in ecology» at the IEE RAS |
| Благодарности Авторы благодарят инженеров ИПЭЭ РАН Ю. А. Горохову и А. С. Чубракову за помощь в оформлении статьи. | Acknowledgement The authors express their gratitude to the engineers of IEE RAS, Yu. A. Gorokhov and A.S. Chubrakov for their assistance in work on this paper. |
| Конфликт интересов Не указан. | Conflict of Interest None declared. |
Список литературы / References
- Бокариус К. И. Экспертиза птичьего пуха / К. И. Бокариус // Практика криминалистической экспертизы. Вып. 1 – Киев, 1961.
- Бордонос Т. Г. О некоторых особенностях строения перьев и пуха домашних животных / Т. Г. Бордонос, Л. П. Булыга // Криминалистическая и судебная экспертиза. Вып. 5. – Киев, 1968.
- Бородулина Т. Л. Строение кроющего оперения птиц в связи с их полётом / Т. Л. Бородулина // Зоол. журн. Т. 43. № 12, 1964. – С. 1826–1836.
- Булыга Л. П. Судебно биологическое исследование перьев и пуха птиц / Л. П. Булыга // Киев: КНИИСЭ, 1998. – 37 с.
- Вараксин А. Н. Метод оценки информативности признаков в задачах обработки многомерных данных / А. Н. Вараксин, Н. И. Куренков, Б. Д. Лебедев // Оборонная техника. №10, 2003. – С. 81–84.
- Добринский Л. Н. Географическая изменчивость варьирования морфологических признаков птиц и некоторые вопросы систематики / Л. Н. Добринский // Внутривидовая изменчивость наземных позвоночных животных и микроэволюция. – Свердловск, 1965. – С. 341–344.
- Ильичёв В. Д. Дополнительные опахала в птерилозисе уха птиц, их строение и функция / В. Д. Ильичёв // ДАН, Т. 144, № 5, 1962. – С. 1185–1188.
- Ильичёв В. Д. Исследование тонкой структуры пера с помощью прибора ФМН 2 / В. Д. Ильичёв // Зоол. журнал, т. 42. № 10, 1963. – С. 1584–1585.
- Силаева О. Л. Определитель птиц по перу и его фрагментам. Отряд Воробьинообразные (Passeriformes). Семейство Врановые (Corvidae) / О. Л. Силаева, В. Д. Ильичёв, О. Ф. Чернова // Саарбрюкен: LAP LAMBERT Acad. Publ., 2012. – 316 с.
- Силаева О. Л. Определитель птиц по перу и его фрагментам. Отряды: Курообразные (Galliformes), Голубеобразные (Columbiformes), Рябкообразные (Pterocliformes) / О. Л. Силаева, В. Д. Ильичёв, О. Ф. Чернова, А. Н. Вараксин // М.: ИПЭЭ, 2013. – 120 с. + CD-ROM.
- Силаева О. Л. Определитель птиц по перу и его фрагментам. Отряд Гусеобразные (Anseriformes) / О. Л. Силаева, О. Ф. Чернова, А. Н. Вараксин // М.: ИПЭЭ, 2015. – 269 c. + CD-ROM.
- Силаева О. Л. Определитель птиц по перу и его фрагментам. Отряд Ржанкообразные (Сharadriiformes) / О. Л. Силаева, О. Ф. Чернова, С. А. Букреев, А. Н. Вараксин // М.: Товарищество научных изданий КМК, 2018. – 385 с.
- Силаева О. Л. Внутривидовая изменчивость микроструктуры покровных перьев кряквы и хохлатой чернети / О. Л. Силаева, А. Н. Вараксин // Международный научно-исследовательский журнал. № 5 (47), ч. 5, 2016. – С. 90−100.
- Чернова О. Ф. Архитектоника перьев и её диагностическое значение / О. Ф. Чернова, В. Ю. Ильяшенко, Т. В. Перфилова // М.: Наука, 2006. – 98 с.
- Яблоков А. В. Изменчивость структур пера и окраски яиц у некоторых птиц / А. В. Яблоков, А. В. Валецкий // Зоологический журнал, т. 51. № 2, 1972. – С. 248–258.
- Brom T. G. Microscopic identification of feather-remains after collisions between birds and aircraft / T. G. Brom // Amsterdam, 1980. – 89 p.
- Brom T. G. Microscopic identification of feathers and feather fragments of Palearctic birds / T. G. Brom // Bijdragen tot de Dierkunde. 56, 1986. – P. 181–204.
- Brom T. G. The diagnostic and phylogenetic significance of feather structures / T. G. Brom // Published Ph.D. thesis, University of Amsterdam, 1991. – 279 p.
- Brom T. G. The Phylogenetic significance of the feather character «flexules» / T. G. Brom, H. Visser // Neth. Zool. V. 39, 1989. – P. 226–245.
- Chandler A. C. A study of the structure of feathers with reference to their taxonomic significance / C. A. Chandler // Univ. Publ. V. 13, 1916. – P. 243–446.
- Dove C. J. Quantification of microscopic feather characters used in the identification of North American plovers / C. J. Dove // Condor. 99. № 1, 1997. – P. 47–57.
- Lowe P. R. On the systematic position of the woodpeckers (Pici), honey-guides (Indicator), hoopoes and others / P. R. Lowe // Ibis. V. 88, 1946. – P. 103−127.
- Lucas A. M. Avian anatomy. Integument / A. M. Lucas, P. R. Stettenheim // Washington: US Dept. Parts 1, 2, 1972 – 750 p.
- May F. H. Ptilology – proposed name for the general study of the plumage of birds / F. H. May // Auk 62, 1945. – p. 308.
- Nitzsch Ch. L. System der Pterylographie / Ch. L. Nitzsch // Halle: Eduard Anton, 1840. – 226 p.
- Prast W. BRIS: A computer based bird remains identification system. Further developments / W. Prast, J. Shamoun, B. Bierhuizen. // Birds of Europe. CD-ROM. – Amsterdam: ETI, 1996.
- Prast W., Shamoun J. Bird Remains Identification System (BRIS) / W. Prast, J. Shamoun // Birds of Europe. CD-ROM. – Amsterdam: ETI, 2001.
Список литературы на английском языке / References in English
- Bokarius K. I. Ekspertiza ptich'ego puha [The expertise of birds’ down]. / K. I. Bokarius // Praktika kriminalisticheskoj ekspertizy. Vyp. 1 – Kiev. – 1961. [in Russian]
- Bordonos T. G. O nekotoryh osobennostyah stroeniya per'ev i puha domashnih zhivotnyh [About of some structure features of feathers and down of domestic animals] / T. G. Bordonos, L. P. Bulyga // Kriminalisticheskaya i sudebnaya ekspertiza. Vyp. 5. – Kiev, 1968. [in Russian]
- Borodulina T. L. Stroenie kroyushchego opereniya ptic v svyazi s ih polyotom [The structure of covering plumage of birds in connection with their flight] / T. L. Borodulina // Zool. zhurn. T. 43. № 12, 1964 – S. 1826–1836. [In Russian]
- Bulyga L. P. Sudebno biologicheskoe issledovanie per'ev i puha ptic [Forensic biological research of birds’ feather and down] / L. P. Bulyga // Kiev: KNIISE, 1998 – 37 s. [in Russian]
- Varaksin A. N. Metod ocenki informativnosti priznakov v zadachah obrabotki mnogomernyh dannyh [Method of informational assessment of signs in problems of multidimensional data processing] / A. N. Varaksin, N. I. Kurenkov, B. D. Lebedev // Oboronnaya tekhnika. №10, 2003 – S. 81–84. [in Russian]
- Dobrinskij L. N. Geograficheskaya izmenchivost' var'irovaniya morfologicheskih priznakov ptic i nekotorye voprosy sistematiki [Geographical variability of morphological characteristics variation in birds and some questions of systematics] / L. N. Dobrinskij // Vnutrividovaya izmenchivost' nazemnyh pozvonochnyh zhivotnyh i mikroevolyuciya. – Sverdlovsk, 1965. – S. 341–344. [in Russian]
- Il'ichyov V. D. Dopolnitel'nye opahala v pterilozise uha ptic, ih stroenie i funkciya [Additional vanes in pterilosis of birds’ ear, their structure and function] / V. D. Il'ichyov // DAN, T. 144, № 5, 1962. – S. 1185–1188. [in Russian]
- Il'ichyov V. D. Issledovanie tonkoj struktury pera s pomoshch'yu pribora FMN 2 [The research of a fine feather structure using the device FMN 2] / V. D. Il'ichyov // Zool. zhurnal, VOL. 42. № 10, 1963. – S. 1584–1585. [in Russian]
- Silaeva O. L. Opredelitel' ptic po peru i ego fragmentam. Otryad Vorob'inoobraznye (Passeriformes). Semejstvo Vranovye (Corvidae) [Birds’ identification guide by feathers and its fragments. Passeriformes, family Corvidae] / O. L. Silaeva, V. D. Il'ichyov, O. F. Chernova // Saarbryuken: LAP LAMBERT Acad. Publ., 2012. – 316 s. [in Russian]
- Silaeva O. L. Opredelitel' ptic po peru i ego fragmentam. Otryady: Kuroobraznye (Galliformes), Golubeobraznye (Columbiformes), Ryabkoobraznye (Pterocliformes) [Birds’ identification guide by feathers and its fragments. Orders: Galliformes, Columbiformes, Pterocletiformes] / O. L. Silaeva, V. D. Il'ichyov, O. F. Chernova, A. N. Varaksin // M.: IPEE, 2013. – 120 s. + CD-ROM. [in Russian]
- Silaeva O. L. Opredelitel' ptic po peru i ego fragmentam. Otryad Guseobraznye (Anseriformes) [Birds’ identification guide by feathers and its fragments. Order Anseriformes)] / O. L. Silaeva, O. F. Chernova, A. N. Varaksin // M.: IPEE, 2015. – 269 c. + CD-ROM. [in Russian]
- Silaeva O. L. Opredelitel' ptic po peru i ego fragmentam. Otryad Rzhankoobraznye (Charadriiformes) [Birds’ identification guide by feathers and its fragments. Order Charadriiformes)] / O. L. Silaeva, O. F. Chernova, S. A. Bukreev, A. N. Varaksin // M.: Tovarishchestvo nauchnyh izdanij KMK, 2018. – 385 s. [in Russian]
- Silaeva O. L. Vnutrividovaya izmenchivost' mikrostruktury pokrovnyh per'ev kryakvy i hohlatoj cherneti [Intraspicific variability of microstructure of cover feathers of Anas plathyrinchos and Aythya fuligula / O. L. Silaeva, A. N. Varaksin // Mezhdunarodnyj nauchno-issledovatel'skij zhurnal. № 5 (47), ch. 5, 2016. – S. 90−100. [in Russian]
- Chernova O. F. Arhitektonika per'ev i eyo diagnosticheskoe znachenie [Feather structure and its diagnostic value] / O. F. Chernova, V. Yu. Il'yashenko, T. V. Perfilova // M.: Nauka, 2006. – 98 s. [in Russian]
- Yablokov A. V. Izmenchivost' struktur pera i okraski yaic u nekotoryh ptic [Variability of feather structure and eggs color in some birds] / A. V. Yablokov, A. V. Valeckij // Zoologicheskij zhurnal, VOL. 51. № 2, 1972. – S. 248–258. [in Russian]
- Brom T. G. Microscopic identification of feather-remains after collisions between birds and aircraft / T. G. Brom // Amsterdam, 1980. – 89 p.
- Brom T. G. Microscopic identification of feathers and feather fragments of Palearctic birds / T. G. Brom // Bijdragen tot de Dierkunde. V. 56, 1986. – P. 181–204.
- Brom T. G. The diagnostic and phylogenetic significance of feather structures / T. G. Brom // Published Ph.D. thesis, University of Amsterdam, 1991. – 279 p.
- Brom T. G. The Phylogenetic significance of the feather character «flexules» / T. G. Brom, H. Visser // Neth. Journ. Zool. V. 39, 1989. – P. 226–245.
- Chandler A. C. A study of the structure of feathers with reference to their taxonomic significance / C. A. Chandler // Univ. Calif. Publ. V. 13, 1916. – P. 243–446.
- Dove C. J. Quantification of microscopic feather characters used in the identification of North American plovers / C. J. Dove // Condor. V. 99. № 1, 1997. – P. 47–57.
- Lowe P. R. On the systematic position of the woodpeckers (Pici), honey-guides (Indicator), hoopoes and others / P. R. Lowe // Ibis. V. 88, 1946. – P. 103−127.
- Lucas A. M. Avian anatomy. Integument / A. M. Lucas, P. R. Stettenheim // Washington: US Dept. Agricult. Parts 1, 2, 1972 – 750 p.
- May F. H. Ptilology – proposed name for the general study of the plumage of birds / F. H. May // Auk 62, 1945. – p. 308.
- Nitzsch Ch. L. System der Pterylographie / Ch. L. Nitzsch // Halle: Eduard Anton, 1840. – 226 p. [In German]
- Prast W. BRIS: A computer based bird remains identification system. Further developments / W. Prast, J. Shamoun, B. Bierhuizen. // Birds of Europe. CD-ROM. – Amsterdam: ETI, 1996.
- Prast W., Shamoun J. Bird Remains Identification System (BRIS) / W. Prast, J. Shamoun // Birds of Europe. CD-ROM. – Amsterdam: ETI, 2001.