ЭЭГ ЗРИТЕЛЬНОЙ И СЕНСОМОТОРНОЙ ОБЛАСТЕЙ КОРЫ ГОЛОВНОГО МОЗГА КРЫСЯТ РАННЕГО ВОЗРАСТА, ПЕРЕНЕСШИХ ВЛИЯНИЕ ГИПОКИНЕЗИИ В ЗАРОДЫШЕВЫЙ ПЕРИОД ПРЕНАТАЛЬНОГО РАЗВИТИЯ
Махмудова Н.Ш.
Институт Физиологии им. А.И. Гараева, НАН Азербайджана, лаборатория Факторы среды и формирования анализаторов
ЭЭГ ЗРИТЕЛЬНОЙ И СЕНСОМОТОРНОЙ ОБЛАСТЕЙ КОРЫ ГОЛОВНОГО МОЗГА КРЫСЯТ РАННЕГО ВОЗРАСТА, ПЕРЕНЕСШИХ ВЛИЯНИЕ ГИПОКИНЕЗИИ В ЗАРОДЫШЕВЫЙ ПЕРИОД ПРЕНАТАЛЬНОГО РАЗВИТИЯ
Аннотация
Изучена динамика формирования биоэлектрической активности зрительной и сенсомоторной зон коры мозга крысят, переживших влияние фактора гипокинезии в ранние периоды эмбриогенеза. Условия гипокинезии беременных крыс создавались в специальных клетках с учетом биометрических критериев экспериментальных животных. Выявлено, что гипокинезия, как неблагоприятный фактор среды, применяемый в ранние периоды эмбриогенеза, способствует нарушению динамики формирования амплитудно-частотных свойств суммарной биоэлектрической активности коры мозга у потомства. При этом наиболее существенные сдвиги в сроках формирования биоэлектрической активности мозга проявляются в диапазоне высокоамплитудных низкочастотных волн ЭЭГ в сенсомоторной коре, а в диапазоне медленных высокочастотных волн ЭЭГ в зрительной коре животных раннего постнатального развития.
Ключевые слова: гипокинезия, онтогенез, электроэнцефалограмма, крысы
Mahmudova N.Sh.
Institute of Physiology named after A.I. Garayev of Azerbaijan National Academy of Sciences, Laboratory Environmental factors and the formation of analyzers
EEG OF VISUAL AND SENSORIMOTOR CEREBRAL CORTEX OF THE INFANT RATS OF EARLY AGE WHO TRANSFERRED INFLUENCE OF A HYPOKINESIA TO THE GERMINAL PERIOD OF PRENATAL DEVELOPMENT
Abstract
Dynamics of formation of bioelectric activity of visual and sensorimotor cerebral cortex of the infant rats who endured influence of a factor of a hypokinesia during the early periods of an embryogenesis is studied. Conditions of a hypokinesia of pregnant rats were created in special cages taking into account biometric criteria of experimental animals. It is revealed that the hypokinesia as the adverse factor of the environment applied during the early periods of an embryogenesis promotes violation of dynamics of formation of amplitude-frequency properties of total bioelectric activity of bark of a brain at posterity. Thus the most essential shifts in terms of formation of bioelectric activity of a brain are shown in the range of high-amplitude low-frequency waves of EEG in sensorimotor cerebral cortex, and in the range of slow high-frequency waves of EEG in visual cerebral cortex of animals of early post-natal development.
Keywords: hypokinesia, ontogenesis, electroencephalogram, rats
Эмбриональный и ранний постэмбриональный периоды онтогенеза являются исключительно важными звеньями с точки зрения исследования динамики развития головного мозга. Не вызывает сомнения то, что развитие и свойства этого органа определяются, прежде всего, генетическими факторами [4,7]. В связи с этим необходимо подчеркнуть некоторые принципы генетического контроля важнейших элементарных процессов, из которых слагается формирование мозга в эмбриональном периоде [11]:
1) дифференцировка части клеток эктодермы в нейроэктодерму,
2) размножение клеток нейроэктодермы и их дифференцировка в нейробласты (предшественники нейронов) и глиобласты (предшественники глиоцитов),
3) миграция в зоны окончательного их нахождения,
4) образование из этих клеток различных типов нейронов и глиальных клеток,
5) установление синаптических связей между нейронами,
6) запрограммированная гибель части клеток.
Несмотря на строгую детерминированность развития перечисленных процессов, среда, создаваемая в комплексе с внешними факторами, играет важное значение для формирование мозга. С учетом сказанных представлялся актуальным вопрос о динамике биоэлектрических показателей мозга в условиях воздействия гипокинезии.
Наша задача была посвящена изучению возрастной динамики формирования биоэлектрической активности зрительной и сенсомоторной областей коры головного мозга, а также изменений в структуре ЭЭГ после влияния внутриутробной гипокинезии. В литературе встречаются аналогичные исследования, охватывающие отдельные аспекты данной проблемы [8,10,13,18]. Тем не менее, данные о динамике основных характеристик ЭЭГ в онтогенезе разрозненны и противоречат друг другу.
По существующим общепринятым правилам пренатальный онтогенез или внутриутробный период развития разделяется на несколько этапов: зародышевый - первые 10-12 дней после образование зиготы, предплодный – 10-15 дней и плодный - 16-21-е дни беременности белых крыс. По сведениям других исследователей возрастная периодизация развития белых лабораторных крыс представлена следующим образом:
10-12 суток (зародышевый), 15 суток (предплодный), 20-21 суток (плодный), новорожденные (сосунки), 1-2 неделя (подсосунки), 1 месяц (инфантильный), 6 месяцев (репродуктивный), 12 месяцев (зрелый) и 2 года (старческий) [5].
В нашем случае мы придерживались первого типа периодизации – условно именуемые как критические периоды эмбрионального онтогенеза белых крыс.
Нормальное развитие мозга может нарушаться под влиянием многих факторов. Это определяется как высокой чувствительностью мозга в критические периоды развития, так и необратимостью некоторых из последствий таких воздействий [6]. Последнее зависит от многих причин, одной из которых является ограничение двигательной активности.
Методика исследований
Крысы линии Вистар - объект наших исследований – являются наиболее распространенной линией крыс, характеризующихся своими генетическими особенностями. Размеры тела взрослых крыс 150-250 см, вес 150-230 г. Продолжительность жизни 3-4 года, из которых около 2-х лет крысы активно размножаются. Продолжительность беременности варьирует от 20 до 26 дней и составляет в среднем около 22 дней. В работе было использовано потомство в количестве 136 (75 контрольных и 46 экспериментальных) животных, которое получено от 29 (8 контрольных и 21 экспериментальных) крыс. Животные содержались в сухом, оттапливаемом помещении, с хорошим естественным и искусственным освещением и принудительной вентиляцией. Температура в виварии была в пределах 20-220С. Содержались крысы в металлических клетках с размерами 50х30х30 см. Клетка была предназначена для 5 самок [5]. Во избежание незапланированной беременности самки и самцы содержались в различных клетках. В нужное для нас время в каждую клетки с самками подсаживали по одному самцу. Определение беременности устанавливалось путем взятия мазка. После определения беременности все самки были разделены на 2 большие группы: контрольную и экспериментальную. Контрольная группа животных продолжала содержаться в прежних обычных условиях. В зависимости от того, на какой период беременности мы хотим воздействовать, животные помещались в условия гипокинезии. Для создания условия гипокинезии нами были сконструированы специальные клетки размером 14х8х20 см, в которые помещались беременные самки по одной особи в каждую. Поилка и кормушка располагались снаружи и были жестко прикреплены к клетке. Гипокинезия беременных самок крыс создавалась в зародышевый (Е0-Е7) период онтогенеза, а остальные сроки беременности животные размещались в обычных условиях вивария.
Все полученное потомство как контрольных, так и экспериментальных животных, было разделено на 2 подгруппы:
1) контрольные крысята всех возрастных групп, развившиеся в нормальных условиях в качестве общего контроля;
2) 10-ти, 20-ти, 30-ти дневные крысята, развившиеся в зародышевый период антенатального онтогенеза в условиях ограничения подвижности матерей.
Регистрация и анализ биоэлектрической активности с поверхности зрительной и сенсомоторной областей коры головного мозга животных была произведена с применением компьютерного 18-ти канального энцефаллографа «Нейрон Спектр-2» фирмы «Нейрософт». Статистическая обработка экспериментальных данных производилась при помощи пакета программ Statistical for Windows [3]. Эксперименты проводились с соблюдением принципов «Европейской конвенции о защите позвоночных животных, которые используются для экспериментальных и других научных целей» [Страсбург, 1986] и постановления первого национального конгресса по биоэтике [Киев, 2001]. Для оценки динамики проявления частотного состава биоэлектрической активности зрительной коры мозга крысят раннего возраста, были анализированы 10 секундные стабильные отрезки нативной записи ЭЭГ как правого, так и левого полушарного отведения.
Полученные результаты и их обсуждения
Результаты спектрального анализа биоэлектрической активности правого полушария зрительной коры головного мозга контрольных животных показал, что количество дельта волн – преимущественной частотой 2-4 кол/сек с возрастом увеличивается с 1,6% у 10-ти дневных до 4,7% у 20-ти дневных. В то время как у 30-ти дневных животных наблюдается уменьшение 2,6%. Выявлено, что у бодрствующих контрольных крысят в спектре ЭЭГ волны тета диапазона незначительно колеблются и противоположно предыдущим в начале незначительно уменьшаются с 27,7% у 10-ти дневных и 27,6% у 20-ти дневные до 25,5% у 30-ти дн. Представленность в спектре ЭЭГ волн альфа диапазона с возрастом увеличивается, достигая 27,5% у 20-ти дневных и незначительным превалированием у 30-ти дневных. Обнаружено, что спектр бета диапазона ЭЭГ со становлением возраста уменьшается с 49,6% у 10-ти дневных до 40,2% у 20-ти дневных, далее у 30-ти дневных наблюдается увеличение количество волн до 45,3%.
Анализ ЭЭГ правого полушария зрительной коры головного мозга животных, подверженных гипокинезии в зародышевый период эмбрионального развития, выявил, что как и у нормально развившихся, у потомства экспериментальных животных, регистрируется полный спектр биоэлектрической активности коры мозга. Вместе с тем установлено, что по мере взросления животных представленность дельта волн в спектре ЭЭГ снижается. Как видно из диаграммы 1 возрастные различия проявления тета волн у 10-ти и 20-ти дневных животных почти незначительны и приблизительно равны 21% от общего спектра ЭЭГ. Однако к 30-ти дневному возрасту этот показатель увеличивается до 25,8. Проявление альфа волн с возрастом животных увеличивается с 31,7% у 10-ти дневных до 33,5% у 20-ти дневных крысят. При этом интенсивность проявления альфа ритма ЭЭГ уменьшается к 30-ти дневному возрасту (30,4%). Динамика бета волн спектра ЭЭГ у отдельных экспериментальных групп животных в основном схожая и выражена почти в одинаковой степени за исключением животных 20-ти дневного возраста. В этой группе животных встречаемость бета волн в спектре ЭЭГ составляла 43% от общего количества колебаний. Подробные данные спектрального анализа ЭЭГ приведены в рис.1.
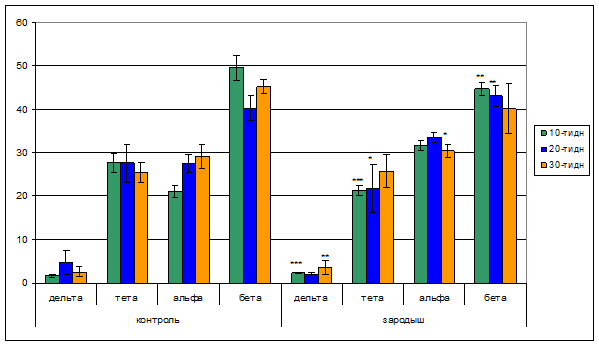
Рис.1. Диаграммы спектрального анализа ЭЭГ правого полушария зрительной коры головного мозга животных, подверженных гипокинезии в зародышевый период эмбрионального развития.
Спектральный анализ ЭЭГ левого полушария зрительной коры головного мозга контрольных животных показал, что представленность дельта волн спектра биоэлектрической активности как у 10-ти, так и у 20-ти дневных животных приблизительно одинаковая и равна 3,4-3,2% соответственно. В то время как у 30-ти дневных происходит существенное увеличение до 7,6%. Оказалось, что представленность дельта диапазона с преимущественной частотой 2,5-4 Гц составляет всего 2,76 (10.8%) от общего составляющегося спектра ЭЭГ. Более иная картина наблюдается в динамике высокочастотных составляющих спектра ЭЭГ. Обращает на себя внимание то, что со становлением возраста представленность альфа диапазона возрастает и составляет приблизительно 38-39% спектра ЭЭГ у 20-ти и 30-ти дневных животных. Возможно, довольно высокий уровень представленности альфа ритма в спектре ЭЭГ в ранние периоды развития связан с наличием так называемых альфа подобных волн ЭЭГ, отмеченные в некоторых литературных источниках [14]. Приблизительно близкие значения и похожая динамика наблюдается в бета диапазоне ЭЭГ животных. Здесь мы так же наблюдаем, что в первый месяц (10,20,30 дневные возрасты) постнатальной жизни бета волны составляет 36-41% общего диапазона ЭЭГ.
Аналогичная картина распределения волн ЭЭГ была наблюдена при анализе биоэлектрической активности из левого полушария мозга экспериментальных животных. Результаты анализа показали, что количество дельта волн по мере становления возраста существенно увеличиваясь и затем уменьшаясь, достигает 2,2% у 30-ти дневных. Тета волны спектра ЭЭГ с возрастом незначительно уменьшаются с 22,4% у 10-ти дневных до 21,3% у 20-ти дневных, а затем вновь увеличиваются до 25,4% у 30-ти дневных. Процентное соотношение альфа диапазона с возрастом уменьшается с 39,0% у 10-ти дневных животных до 32,4% у 20-ти дневных с дальнейшим увеличением этого показателя у 30-ти дневных крысят до 35,1%. Волны бета диапазона ЭЭГ экспериментальных животных с возрастом увеличиваются с 36,9% у 10-ти дневных до 41,0% и 37,3% у 20-ти и 30-ти дневных соответственно (рис.2).
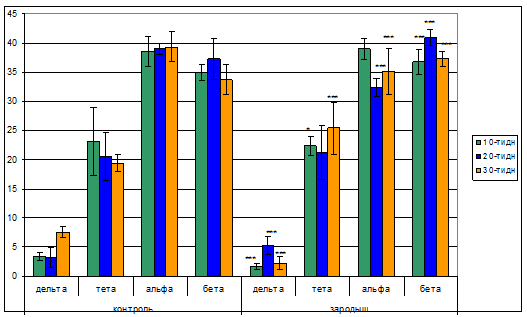
Рис.2. Диаграммы спектрального анализа ЭЭГ левого полушария зрительной коры головного мозга животных, подверженных гипокинезии в зародышевый период эмбрионального развития
Спектральный анализ ЭЭГ сенсомоторной коры головного мозга показал, что по сравнению со зрительной корой степень представленности всего диапазона ЭЭГ сенсомоторной области коры существенно увеличивается. В тоже время было выявлено, что отдельные частотно-составляющее компоненты ЭЭГ активности по-разному представлены и зависят от возраста животных. Так оказалось, что дельта волны с возрастом уменьшаются с 7,7% до 3,2% у 10-ти и 20-ти дневных крысят, а затем увеличиваются до 4,9% у 30-ти дневных. Схожие изменения в динамике тета диапазона ЭЭГ выявлены и в сенсомоторной коре контрольных животных. При этом представленность тета волн в спектре ЭЭГ составлял 42,7% у 10-ти дневных, 31,1% у 20-ти, и 28,2% у 30-ти дневных. Количество волн альфа диапазона с возрастом увеличивается с 25,9% у 10-ти до 31,3% и 38,5% 20-ти и 30-ти дней соответственно. Несмотря на то, волны бета диапазона с возрастом увеличиваются с 23,7% у 10-ти дневных до 34,4% у 20-ти дневных крысят, затем вновь уменьшаются до 28,4% у 30-ти дневных.
Статистический анализ биоэлектрической активности сенсомоторной области коры головного мозга показал, что у животных 10-ти дневного постнатального развития уже регистрируется полноценное ЭЭГ. Вместе с тем условия пренатальной гипокинезии способствуют появлению некоторых различий, наблюдаемых у контрольных животных. Так, проведенный анализ ЭЭГ правого полушария сенсомоторной коры головного мозга животных, подверженных гипокинезии в зародышевый период эмбрионального развития показал, что с возрастом количество дельта волн уменьшается с 6,5% у 10-ти дневных до 2,4% и 1,5% у 20-ти и 30-ти дневного возраста соответственно. Выявлено, что тета волны ЭЭГ значительно меньше представлены у 20-ти дневных крысят, (см.рис. 3) вместе с тем с увеличением возраста животных в 30 дней возрастает до 25,3%. Иная картина наблюдалась динамике альфа ритмов ЭЭГ. Обнаружено, что волны альфа диапазона, с возрастом незначительно уменьшаясь к 20-ти дневному возрасту, затем увеличиваются до 36,1% у 30-ти дней. При этом какие-либо упорядоченные колебание волны бета диапазона не наблюдаются.Данные анализа ЭЭГ сенсомоторной коры у животных подробно представлены в рис. 3.
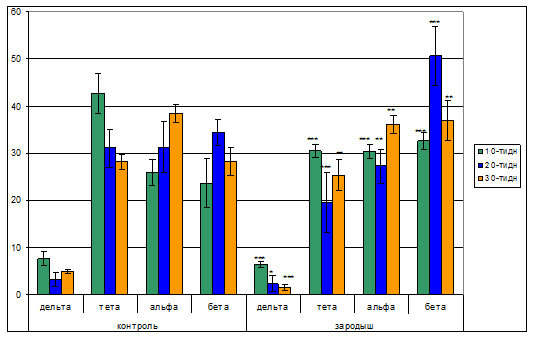
Рис.3. Диаграммы спектрального анализа ЭЭГ правого полушария сенсомоторной коры головного мозга животных, подверженных гипокинезии в зародышевый период эмбрионального развития
Проводимый нами анализ показывает, что как и при правополушарном отведении ЭЭГ левого полушария сенсомоторной коры головного мозга контрольных животных характеризуются однонаправленными изменениями частотных показателей. Так, выявлено, что в спектре ЭЭГ дельта волны с возрастом уменьшаются с 5,9% у 10-ти дневных до 3,4% и 1,2% у 20-ти и 30-ти дневных соответственно. Волны тета диапазона ЭЭГ колеблются в пределах 29,5% и 22% у 10-ти дневных и 30-ти дневных соответственно. Количество волн альфа диапазона незначительно колеблются в пределах 34,1% и 40,1% у 10-ти дневных и у 30-ти дневных соответственно. Волны бета диапазона с возрастом увеличиваются с 30,5% у 10-ти дневных до 38,4% у 20-ти дневных, с дальнейшим уменьшением у 30-ти дневных – 36,7%. На рис. 4 показаны данные спектрального анализа ЭЭГ левого полушария сенсомоторной коры головного мозга животных, подверженных гипокинезии в зародышевый период эмбрионального развития. Обращает на себя внимание тот факт, что дельта волн спектра ЭЭГ наиболее заметно представлены у животных, находящихся в 3-х и 4-х недельном возрасте постнатального развития. При этом представленность волн дельта диапазона ЭЭГ составляет в среднем 3-3,5%. В дальнейшем у 3-х месячных животных происходит уменьшение ее выраженности до 2,1%. Представленность в спектре ЭЭГ тета волн у экспериментальных групп животных распределились в следующем порядке: у 10-ти дневных – 30,8%, у 20-ти дневных 15,7%, у месячных – 25,4%. Выявлено, что представленность альфа ритмов ЭЭГ в первой декаде постнатального возраста существенно с возрастом уменьшается от 31,4% до 27,4% у 20-ти дневных, затем увеличивается до 28,3% у 30-ти дневных. Встречаемость бета волн спектра ЭЭГ у экспериментальных групп животных варьирует в пределах 42-55%, за исключением ранней декады (10-ти дневных), где этот ритм был выявлен в 36,7% (рис.4).
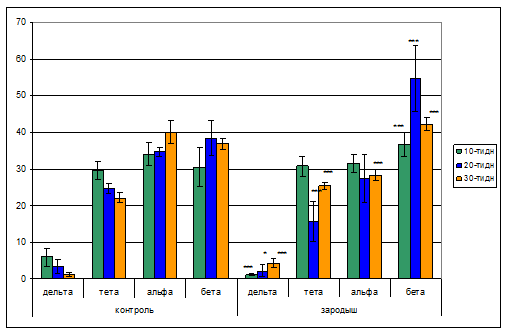
Рис.4. Диаграммы спектрального анализа ЭЭГ левого полушария сенсомоторной коры головного мозга животных, подверженных гипокинезии в зародышевый период эмбрионального развития
Существующие экспериментальные данные подтверждают возможность регистрации биоэлектрической активности мозга у животных в поздние периоды пренатального онтогенеза, сходные по некоторым параметрам к дефинитивному. Вместе с тем, известно, что формирование активности основных ритмов ЭЭГ, в основном, происходит в первый месяц постнатального онтогенеза животных. По данным многих исследователей среди признаков зрелости в динамике биоэлектрической активности головного мозга важную информацию носит частотно-амплитудный спектр ЭЭГ. При этом особое внимание уделяется альфа-ритмам, который постепенно и длительно формируется с возрастом [1,12,15].
Исследованиями подтвержден факт об опережающем значении созревания затылочных областей коры, чем лобных [16]. Вероятно, это объясняется опережающем морфофункциональным созреванием их нисходящих связей по сравнении с другими структурами мозга [9], Становление и полноценное проявление биоэлектрической активности мозга связано с созреванием коры головного мозга, и в первую очередь с формированием структур интернейронов, создающих условия для периодичности активационных и тормозных процессов [13].
Анализ особенностей центральной регуляции репродуктивной функции у женщин при беременности в норме и при осложненных формах ее протекания, дает возможность предположить о нарушениях взаимоотношений гипоталамо-гипофизарных структурах, так как именно они в этот период испытывают наибольшую дополнительную нагрузку в своей работе [17]. Косвенную оценку таких функциональных изменений их активности можно осуществить при анализе ЭЭГ [2], которая в достаточной мере чувствительна к функциональному состоянию головного мозга [19].
Таким образом, результаты проведенных исследований позволяют прийти к следующим выводам:
- гипокинезия как неблагоприятный фактор среды, применяемый в ранние периоды эмбриогенеза, способствует нарушению динамики формирования амплитудно-частотных свойств суммарной биоэлектрической активности коры мозга у потомства.
- наиболее существенные сдвиги в сроках формирования биоэлектрической активности мозга проявляются в диапазоне высокоамплитудных низкочастотных волн ЭЭГ в сенсомоторной коре, а в диапазоне медленных высокочастотных волн ЭЭГ в зрительной коре животных раннего постнатального развития.
Литература
- Безруких М.М., Мачинская Р.И., Фарбер Д.А. Мозговая организация когнитивных процессов в дошкольном возрасте //Пед. вестник. М.: Академия.- 2009.
- Болдырева Г.Н., Шарова Е.В., Добронравова И.С."Роль регуляторных структур в формировании ЭЭГ человека" // ж. Физиология человека.- 2000.- №.- С.19-34.
- Гублер Е.А., Генкин А.А. Применение непараметрических критериев статистики в медико-биологических исследованиях.- 1973.- С.123.
- Жимулев И.Ф. Генетическая детерминированность поведения дрозофилы и человека // Соросовский образовательный журнал.- 2000.- т.6, № 1.-С. 22-25.
- Западнюк И.П., Захария Е.А.Лабораторные животные. Разведение, содержание, использование в эксперименте. Кн.: 3-е изд., перераб. и доп. Киев: Вища школа.- 1983.-С.- 383.
- Кассиль В. Г., Отеллин В. А., Хожай Л. И. Критические периоды развития головного мозга // Рос. физиол. ж. им. И. М.Сеченова.- 2000.- Т.86, № 11.- С. 1418— 1425.
- Корочкин Л.И. Гены и поведение. //Соросовский образовательный журнал . – 1997.- № 1.-С.15-21.
- Королева Н.В.,. Небера С.А, Гутник И.Н. Ведущие показатели зрелости биоэлектрической активности головного мозга у детей в возрасте от 1 до 7 лет // Физиология человека. –2002. – Т. 28, № 6. – С. 57–63.
- Кулаковский Ю.В., Дубровинская Н.В. Возрастные особенности мозговой организации вербальной деятельности: электрофизиологический анализ // Физиология человека. – 1997. – Т. 23, № 3. – С. 122–124.
- Новикова Л.А. Возрастные особенности электрической активности мозга детей и подростков // Журн. высш. нерв. деят. – 1961. – Т.11, вып. 1. С. 60–70.
- Рыжавский Б.Я. Развитие головного мозга в ранние периоды онтогенеза // Соровский образовательный журнал. 2000.- т.6, №1.- С.37-43.
- Строганова Т.А., Дегтярева М.Г, Володин Н.Н. Электроэнцефалография в неонатологии. Вестник интенсивной терапии. - №1.-С. 60—63.
- Фарбер Д.А. Вильдавский В.Ю. Гетерогенность и возрастная динамика α-ритма электроэнцефалограммы / // Физиология человека. – 1996. – Т. 22, № 5. – С. 5–12.
- Фефилов А.В. "Возрастные особенности частотно-специфических характеристик ЭЭГ"., автореферат канд.диссер.,Москва, 2003.
- Цехмистренко Т.А., Черных Н.А., Шеховцев И.К. Постнатальные преобразования микроструктуры //Физиология человека. –т.36, №1.
- Gasser T, Jennen-Steinmetz C, Sroka L, Verleger R, Mocks J. Development of the EEG of school-age children and adolescents. II. Topography. Electroencephalography and Clinical Neurophysiology.1988;69:100–109
- Jan M, Destrieux C. Pituitary disorders in pregnancy.Neurochirurgie. 2000;46(2):88–94.
- Johnstone S.J., Barry,J R.J., Anderson W. et al. Age-related changes in child and adolescent event-related potential component morphology, amplitude and latency to standard and target stimuli in an auditory oddball task / // Int J Psychophysiol. – 1996. – V. 24, № 3. – Р. 223–238.
- Van den Bergh BRH, Mulder EJH, Mennes M, Glover V. Antenatal maternal anxiety and stress and the neurobehavioral development of the fetus and child: links and possible mechanisms. A review. Neurosci Biobehav,2005, Rev 29: 237-258.
References
- Bezrukih M.M., Machinskaja I., Farber D.A. Mozgovaja organizacija kognitivnyh processov v doshkol'nom vozraste //Ped. vestnik. M.: Akademija.- 2009.
- Boldyreva G.N., Sharova E.V., Dobronravova I.S. "Rol' reguljatornyh struktur v formirovanii JeJeG cheloveka" // zh. Fiziologija cheloveka.- 2000.- №.- S.19-34.
- Gubler E.A., Genkin A.A. Primenenie neparametricheskih kriteriev statistiki v mediko-biologicheskih issledovanijah.- 1973.- S.123.
- Zhimulev I.F. Geneticheskaja determinirovannost' povedenija drozofily i cheloveka // Sorosovskij obrazovatel'nyj zhurnal.- 2000.- t.6, № 1.-S. 22-25.
- Zapadnjuk I.P., Zaharija E.A. Laboratornye zhivotnye. Razvedenie, soderzhanie, ispol'zovanie v jeksperimente. Kn.: 3-e izd., pererab. i dop. Kiev: Vishha shkola.- 1983.-S.- 383.
- Kassil' V. G., Otellin V. A., Hozhaj L. I. Kriticheskie periody razvitija golovnogo mozga // Ros. fiziol. zh. im. I. M.Sechenova.- 2000.- T.86, № 11.- S. 1418— 1425.
- Korochkin L.I. Geny i povedenie. //Sorosovskij obrazovatel'nyj zhurnal . – 1997.- № 1.-S.15-21.
- Koroleva N.V.,. Nebera S.A, Gutnik I.N. Vedushhie pokazateli zrelosti biojelektricheskoj aktivnosti golovnogo mozga u detej v vozraste ot 1 do 7 let // Fiziologija cheloveka. –2002. – T. 28, № 6. – S. 57–63.
- Kulakovskij Ju.V., Dubrovinskaja N.V. Vozrastnye osobennosti mozgovoj organizacii verbal'noj dejatel'nosti: jelektrofiziologicheskij analiz // Fiziologija cheloveka. – 1997. – T. 23, № 3. – S. 122–124.
- Novikova L.A. Vozrastnye osobennosti jelektricheskoj aktivnosti mozga detej i podrostkov // Zhurn. vyssh. nerv. dejat. – 1961. – T.11, vyp. 1. S. 60–70.
- Ryzhavskij B.Ja. Razvitie golovnogo mozga v rannie periody ontogeneza // Sorovskij obrazovatel'nyj zhurnal. 2000.- t.6, №1.- S.37-43.
- Stroganova T.A., Degtjareva M.G, Volodin N.N. Jelektrojencefalografija v neonatologii. Vestnik intensivnoj terapii. 2005.- №1.-S. 60—63.
- Farber D.A. Vil'davskij V.Ju. Geterogennost' i vozrastnaja dinamika α-ritma jelektrojencefalogrammy / // Fiziologija cheloveka. – 1996. – T. 22, № 5. – S. 5–12.
- Fefilov A.V. "Vozrastnye osobennosti chastotno-specificheskih harakteristik JeJeG"., avtoreferat kand.disser.,Moskva, 2003.
- Cehmistrenko T.A., Chernyh N.A., Shehovcev I.K. Postnatal'nye preobrazovanija mikrostruktury // Fiziologija cheloveka. 2010.– t.36, №1.