ИЗМЕНЧИВОСТЬ МОРФОЛОГИИ ЛИСТОВЫХ ПЛАСТИНОК ПРИ СЕМЕННОЙ РЕПРОДУКЦИИ ИВЫ ТРЕХТЫЧИНКОВОЙ
DOI: https://doi.org/10.23670/IRJ.2018.75.9.025
ИЗМЕНЧИВОСТЬ МОРФОЛОГИИ ЛИСТОВЫХ ПЛАСТИНОК ПРИ СЕМЕННОЙ РЕПРОДУКЦИИ ИВЫ ТРЕХТЫЧИНКОВОЙ
Научная статья
Афонин А.А.*
ORCID: 0000-0002-9392-2527,
Брянский государственный университет имени академика И.Г. Петровского, Брянск, Россия
* Корреспондирующий автор (afonin.salix[at]gmail.com)
Аннотация
Изложена история развития взглядов на возможность апомиксиса у видов с различным уровнем плоидности и современное состояние проблемы. Проанализированы проблемы цитогенетики ивы трехтычинковой (Salix triandra L.). Сформулирована цель исследования: обосновать возможность редуцированного гаметофитного апомиксиса при семенной репродукции ивы трехтычинковой и выявить закономерности расщепления по морфологии листовых пластинок при апомиктической репродукции этого вида. Для получения и обработки материала использованы основные методы морфологии растений: экспериментальный, описательный и сравнительный. В семенном потомстве коротколистной формы ивы трехтычинковой, полученном в условиях беспыльцевого режима, выявлено расщепление по морфологии листовых пластинок. Наряду с типичной формой (f. typica), выявлены сеянцы с уникальной морфологией листьев: узколистная (f. angustifolia) и коротколистная (f. brevifolia). Обоснована гипотеза о возможности гаметофитного апомиксиса у ивы трехтычинковой в виде редуцированного партеногенеза с сохранением гаплоидности зародышей и спонтанной соматической диплоидизации на постэмбриональных этапах онтогенеза. Подтверждена гипотеза о полиплоидном происхождении ивы трехтычинковой. Полученные результаты рекомендуется использовать при описании формового разнообразия ивы трехтычинковой в природных популяциях, а также при планировании селекционного процесса и оценке полученного материала.
Ключевые слова: ива трехтычинковая, Salix triandra, гаметофитный апомиксис, редуцированный партеногенез, соматическая диплоидизация.
VARIABILITY OF LAMINA MORPHOLOGY IN SEEDS REPRODUCTION OF ALMOND-LEAVED WILLOW
Research article
Afonin A.A.*
ORCID: 0000-0002-9392-2527,
Ivan Petrovsky Bryansk State University, Bryansk, Russia
* Corresponding author (afonin.salix[at]gmail.com)
Abstract
The article outlines the history of the development of views on the possibility of apomixis in species with different levels of ploidy and the current state of the problem. The problems of cytogenetics of almond-leaved willow (Salix triandra L.) are analyzed. The aim of the research is formulated: To justify the possibility of reduced gametophytic apomixis in seed reproduction of an almond-leaved willow and to reveal the patterns of morphological splitting in lamina during apomictic reproduction of this species. The main methods of plant morphology were used to obtain and process the material: Experimental, descriptive and comparative. We identified a splitting by the morphology of lamina in the seed progeny of a short-leaved form of an almond-leaved willow obtained under the conditions of pollen absence. Along with the typical form (f. typica), we identified seedlings with a unique morphology of leaves: Narrow-leaved (f. angustifolia) and short-leaved (f. brevifolia). The hypothesis about the possibility of gametophytic apomixis of an almond-leaved willow in the form of reduced parthenogenesis with preservation of haploid embryos and spontaneous somatic diploidization in postembryonic stages of ontogenesis is substantiated. The hypothesis of the polyploid origin of an almond-leaved willow was confirmed. Obtained results are recommended to be used when describing the form diversity of an almond-leaved willow in natural populations, as well as when planning the selection process and evaluating the material obtained.
Keywords: almond-leaved willow, Salix triandra, gametophytic apomixis, reduced parthenogenesis, somatic diploidization.
Род Ива (Salix L., Salicaceae Mirb.) представляет собой естественную (монофилетическую) группу двудомных листопадных энтомофильных фанерофитов, характеризующуюся уникальным комплексом признаков: симподиальным типом нарастания побегов, очередным листорасположением, соцветиями сережчатого типа, сильно редуцированным околоцветником и мелкими семенами, снабженными пучком волосков [1, С. 9]. Среди ив широко распространены цитогенетические феномены полиплоидии и анеуплоидии [2, С. 165]. С хозяйственной точки зрения, ивы – это исключительно неприхотливые, быстрорастущие древесные растения, которые широко используются для создания защитных и рекреационно-декоративных насаждений, а также энергетических плантаций. Специфическим продуктом ивоводства является прут, используемый для различных видов плетения [3, С. 51].
Многие виды ив издавна введены в культуру и являются традиционным объектом селекции [3, С. 54]. Интенсивная селекция ив основана на межвидовой гибридизации с последующим отбором перспективных клонов [4, С. 19]. Наследование морфологических признаков при отдаленной гибридизации изучается с середины XIX столетия [5, С. 95]. Однако вскоре было установлено, что у многих растений широко распространен партеногенез [6, С. 311]. В дальнейшем H. Winkler [7, С. 2] доказал, что в растительном царстве существует множество типов апомиксиса. Для многих видов Salix апомиксис – обычное явление [8, С. 43]. Поэтому опыление чужеродной пыльцой не всегда приводит к истинной гибридизации, и реальность многих описанных морфогибридов ив может быть поставлена под сомнение [1, С. 64].
Распространенность гаметофитного апомиксиса в популяциях растений с различным уровнем плоидности остается предметом для дискуссий [9, С. 1-309; 10, С. 80]. Ранее [11, С. 43] нами было описано наследование морфологических признаков при редуцированном апомиксисе в семенном потомстве полиплоидной ивы шерстистопобеговой (S. dasyclados Wimm. = S. gmelinii Pall.). Однако существуют цитоэмбриологические доказательства апомиксиса в популяциях диплоидной ивы трехтычинковой (S. triandra L.) [12, С. 723].
Ива трехтычинковая – Salix triandra L. 1753 (S. amygdalina L. 1753) – широко известный представитель ив средней полосы России. С практической точки зрения, ива трехтычинковая – ценный прутяной вид: известно около 70 культиваров, предназначенных для плантационного разведения [13, С. 145]. В системе рода Salix L. 1753 ива трехтычинковая традиционно относится к секции Amygdalinae Koch, 1837 подрода Salix (=Amerina, =Armena) Dum. 1825 [14, С. 104]. Однако данные молекулярно-генетических исследований свидетельствуют об особом таксономическом статусе этого вида [14, С. 947].
В одной из пионерных работ по цитогенетике Salix [15, С. 364] основное хромосомное число S. triandra в разных выборках было определено равным х = 19 (как у большинства Salicaceae) или х = 22. Авторы предположили, что увеличение основного хромосомного числа с 19 до 22 обусловлено фрагментацией крупных хромосом. Эта точка зрения в дальнейшем разделялась и другими исследователями [16, С. 454]. В целом, S. triandra считается диплоидом 2n = 2х = 38, однако в некоторых выборках этого вида были обнаружены триплоиды 2n = 3х = 57 и тетраплоиды 2n = 4х = 88 [17, С. 271], [18, С, 622], [19, С. 56]. Столь высокие хромосомные числа, несомненно, являются производными от меньших чисел [17, С. 282]. Не исключено, что современные виды ив являются аллополиплоидами [16, С. 476].
В настоящем исследовании мы предприняли попытку доказать возможность редуцированного гаметофитного апомиксиса при семенной репродукции ивы трехтычинковой и выявить закономерности расщепления по морфологии листовых пластинок при семенной репродукции этого вида.
Материал и методы
Исходный материал для данного исследования был собран в популяциях ивы трехтычинковой на территории Брянского лесного массива. В дальнейшем полученный материал культивировался в салицетуме Брянского государственного университета на протяжении 2000…2017 гг.
В процессе получения материала и обработки полученных экспериментальных данных использовались основные методы морфологии растений: экспериментальный, описательный и сравнительный.
В качестве исходного генетического материала использовалась семья F0, выращенная из семян, полученных в 2000 г. от типичной особи S. triandra. Место сбора: зеленая зона г. Брянска, терраса р. Десны (53.201025 N, 34.507728 E). Условия произрастания: ТЛУ С5, периферия ассоциации Salicetum cinereae Zólyomi 1931 (Alnetea glutinosae Br.-Bl. et R. Tx. ex Westhoff et al. 1946; Salicetalia auritae Doing ex Krausch 1968; Salicion cinereae Müller et Görs ex Passarge 1961). Наиболее продуктивные особи были размножены черенками. В 2010 г. рамета одной из пестичных особей данной семьи в результате свободного внутрисемейного опыления дала семенное потомство F1 с заметным расщеплением по морфологии листьев. Одна из особей F1 характеризовалась короткими листовыми пластинками. Ранее [20, С. 32] эта особь была отнесена нами к коротколистной форме (S. triandra f. brevifolia).
В качестве контроля использовался генетический материал пестичной особи исходного вегетативного поколения R0. Данная особь была выращена из черенка, срезанного в 2009 г. с типичной особи S. triandra. Место сбора: зеленая зона г. Брянска, пойма р. Десны (53,255248 N, 34,382311 E). Условия произрастания: ТЛУ В3, Salicetum triandro-viminalis (Tx. 1931) Lohm. 1952 (Salicetea purpureae Moor 1958; Salicetalia purpureae Moor 1958; Salicion albae Tx. 1955). Рассматриваемая особь характеризуется длинными узкими листьями (S. triandra f. longifolia) [20, С. 32].
В данном исследовании использовались сеянцы F2, выращенные из семян, полученных от коротколистной особи (S. triandra f. brevifolia) в 2017 г. в условиях беспыльцевого режима. Посев семян производился 05.06.2017 на влажную фильтровальную бумагу в чашки Петри, которые накрывались крышками. Проращивание семян велось на рассеянном дневном свету. После того, как главные корни достигли достаточно большой длины (5…7 мм), наиболее сильные всходы в количестве 100 шт. были распикированы по две штуки в пластмассовые стаканчики объемом 0,5 л. В процессе пикировки все сеянцы были поделены на две равные серии (по 25 шт.) для того, чтобы можно было учесть возможное влияние условий выращивания на морфологию листьев. Сеянцы первой серии выращивались на торфо-минеральном субстрате, а сеянцы второй серии – на минеральном. Первоначально все сеянцы выращивались на рассеянном дневном свету под крышками от чашек Петри, и только после появления 3…4 настоящих листьев крышки постепенно снимались, и стаканчики выставлялись на солнечный свет. Одновременно выбраковывались слабые сеянцы: в каждом стаканчике из двух сеянцев оставлялся один.
Все сеянцы на протяжении вегетационного периода 2017 г. выращивались в условиях защищенного грунта. Уход за ними заключался в поливе, опрыскивании водой, профилактической обработке инсектоакарицидами и подкормке жидким комплексным удобрением. В период интенсивного роста регулярно выбраковывались слабые сеянцы, отстающие в развитии. В итоге было сохранено 14 сеянцев из первой серии (tr 01…14) и 10 сеянцев из второй серии (tr 15…24).
Первое описание сеянцев производилось 15.08.2017: с точностью ±0,5 см измерялась их высота h. После этого верхушки были прищипнуты, а затем (по мере отрастания боковых побегов) они вновь прищипывались. Второе описание сеянцев производилось 15.09.2017, когда их нарастание в длину практически завершилось. С точностью ±0,5 мм измерялись размеры листовой пластинки: ширина X и длина Y. Для измерения срезалось по два наиболее крупных листа из средней части побегов. Эти же листья гербаризировались, а затем сканировались.
Обработка полученных результатов заключалась в вычислении индекса продолговатости листовой пластинки X/Y и ранжировании сеянцев каждой серии в порядке возрастания этого индекса. Для визуального анализа полученных результатов использовалась диаграмма рассеяния в системе координат X, Y.
Результаты и обсуждение
От контрольной узколистной особи (S. triandra f. longifolia) в 2013 г. в условиях беспыльцевого режима было получено небольшое количество семян (первое семенное поколение F1) с низкой всхожестью. Сеянцы F1 развивались слабо, и в итоге до высадки в грунт было выращено лишь 5 особей, одна из которых характеризовалась длинными узкими листьями, подобно материнской особи. Среди этих пяти особей две оказались тычиночными, а одна пестичной. Выявленный факт расщепления по морфологии листовых пластинок и по гендерной принадлежности указывает на возможность гаметофитного апомиксиса в виде редуцированного партеногенеза. Установленная возможность апомиксиса позволила перейти к изучению изменчивости морфологических признаков при данном способе семенной репродукции.
Апомиктическое потомство F2 коротколистной особи (S. triandra f. brevifolia) выращивалось в условиях тщательного ухода, что позволило повысить число выживших полноценных особей.
Семена F2 оказались очень мелкими, визуально меньше, чем обычные семена ивы трехтычинковой. Всхожесть собранных семян на фильтровальной бумаге составила практически 100%. Через 10 дней после посева (15.06.207) всходы были распикированы в грунт. Дальнейшее развитие сеянцев протекало крайне медленно, и только к середине июля они перешли в фазу 5…7 настоящих листьев. Однако с конца июля начался бурный рост сеянцев. К середине августа выявились различия между сеянцами по высоте. Сеянцы высотой менее 25 см были выбракованы. Среди сохраненных сеянцев их высота варьировалась в достаточно широких пределах. Например, в первой серии сеянцев (на торфо-минеральном субстрате) был получен следующий ранжированный ряд по высоте (см): 29 – 35 – 41 – 42 – 48 – 56 – 56 – 58 – 58 – 62 – 62 – 64 – 68 – 74. В данном случае первые пять сеянцев оказались сравнительно слабыми, а остальные девять – достаточно сильными.
Когда интенсивное нарастание побегов прекратилось, на каждом сеянце из обеих серий было произведено измерение двух листовых пластинок: их ширины (X, мм) и длины (Y, мм). На основании полученных данных была построена соответствующая диаграмма рассеяния (рис. 1).
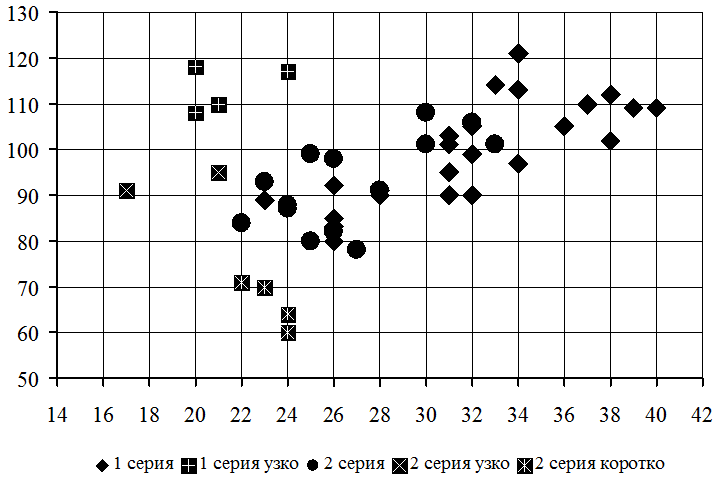
Рис. 1 – Совместная изменчивость ширины и длины листовой пластинки
Примечание: По оси абсцисс ширина листовой пластинки, мм; по оси ординат длина листовой пластинки, мм. В легенде указаны серии и типы конфигурации листовых пластинок: «узко» – узколистные, «коротко» – коротколистные
В общем массиве данных отчетливо выделяется главная последовательность листьев с типичной конфигурацией листовых пластинок: индекс продолговатости варьируется от X/Y = 2,7 до X/Y = 3,9 в первой серии и от X/Y = 2,9 до X/Y = 4,0 во второй серии. Эта последовательность включает два кластера сеянцев: мелколистные (X < 28 мм, Y < 100 мм) и крупнолистные (X > 30 мм, Y > 90 мм). Оба кластера несколько перекрываются по длине листовой пластинки, но хорошо обособлены по ее ширине.
В кластере с шириной листовой пластинки менее 28 мм выявлены уникальные сеянцы. В первой серии два сеянца (tr 13 и tr 14) характеризуются узкими листьями (X/Y = 4,9…5,9). Во второй серии один сеянец (tr 24) характеризуется узкими листьями (X/Y = 4,5…5,4), а два сеянца (tr 15 и tr 16) – короткими листьями (X/Y = 2,5…3,2). На этом основании были выделены три формы, различающиеся по конфигурации листовых пластинок: типичная (f. typica), узколистная (f. angustifolia) и коротколистная (f. brevifolia). Цифровые данные по изменчивости ширины и длины листовых пластинок всех трех форм приведены в таблице (табл. 1).
Таблица 1 – Изменчивость морфологии листовых пластинок
| 1 серия | 2 серия | ||||||||
| № | X, мм | Y, мм | № | X, мм | Y, мм | ||||
| тип. | узкие | тип. | узкие | корот. | |||||
| 1 | 31 | 95 | – | 15 | 24 | – | – | 60 | |
| 1 | 31 | 101 | – | 15 | 24 | – | – | 64 | |
| 2 | 28 | 90 | – | 16 | 23 | – | – | 70 | |
| 2 | 28 | 91 | – | 16 | 22 | – | – | 71 | |
| 3 | 31 | 90 | – | 17 | 27 | 78 | – | – | |
| 3 | 32 | 90 | – | 17 | 26 | 82 | – | – | |
| 4 | 26 | 80 | – | 18 | 33 | 101 | – | – | |
| 4 | 26 | 85 | – | 18 | 32 | 106 | – | – | |
| 5 | 32 | 105 | – | 19 | 25 | 80 | – | – | |
| 5 | 36 | 105 | – | 19 | 24 | 87 | – | – | |
| 6 | 26 | 83 | – | 20 | 28 | 91 | – | – | |
| 6 | 34 | 97 | – | 20 | 23 | 93 | – | – | |
| 7 | 23 | 89 | – | 21 | 30 | 101 | – | – | |
| 7 | 26 | 92 | – | 21 | 30 | 108 | – | – | |
| 8 | 32 | 99 | – | 22 | 24 | 88 | – | – | |
| 8 | 37 | 110 | – | 22 | 22 | 84 | – | – | |
| 9 | 38 | 112 | – | 23 | 26 | 98 | – | – | |
| 9 | 39 | 109 | – | 23 | 25 | 99 | – | – | |
| 10 | 33 | 114 | – | 24 | 21 | – | 95 | – | |
| 10 | 34 | 113 | – | 24 | 17 | – | 91 | – | |
| 11 | 38 | 102 | – | ||||||
| 11 | 40 | 109 | – | ||||||
| 12 | 31 | 103 | – | ||||||
| 12 | 34 | 121 | – | ||||||
| 13 | 24 | – | 117 | ||||||
| 13 | 20 | – | 108 | ||||||
| 14 | 21 | – | 110 | ||||||
| 14 | 20 | – | 118 | ||||||
Примечание: X, мм – ширина листовой пластинки, Y, мм – длина листовой пластинки, «тип.» – типичная форма, «узкие» – узколистная форма, «корот.» – коротоколистная форма.
Полученные формы могут быть выявлены и визуально. На рисунке 2 показаны листовые пластинки узколистной и типичной форм в первой серии, а на рисунке 3 – листовые пластинки узколистной, типичной и коротколистной форм во второй серии (масштабная линейка 100 мм).

Рис. 2 – Формы листовой пластинки в первой серии сеянцев
Примечание: Указаны номера сеянцев. Цена деления масштабной линейки 1 см.

Рис. 3 – Формы листовой пластинки во второй серии сеянцев
Примечание: Указаны номера сеянцев. Цена деления масштабной линейки 1 см.
Выводы
Замедленное развитие сеянцев, выращенных из семян, полученных в условиях беспыльцевого режима, на ранних этапах онтогенеза (в ювенильном возрастном состоянии) указывает на возможность гаметофитного апомиксиса в виде редуцированного партеногенеза с сохранением гаплоидности зародышей. Бурное развитие этих же сеянцев на последующих этапах онтогенеза (в имматурном возрастном состоянии) указывает на возможность спонтанной соматической диплоидизации.
В семенном потомстве коротколистной формы ивы трехтычинковой наблюдается расщепление по морфологии листовых пластинок. Большинство выращенных сеянцев характеризуется типичной конфигурацией листовых пластинок (f. typica). Некоторые сеянцы сохранили признаки коротколистной формы (f. brevifolia) подобно материнской особи. Однако некоторые сеянцы характеризуются узкими листовыми пластинками (f. angustifolia). Узколистная форма выявлена нами впервые (в отличие от сравнительно часто встречающейся длиннолистной формы f. longifolia). Само наличие расщепления дополнительно свидетельствует о возможности гаметофитного апомиксиса в виде редуцированного партеногенеза.
Возможность гаметофитного апомиксиса подтверждает гипотезу о полиплоидном происхождении ивы трехтычинковой. Выщепление типичных, а также узколистных форм указывает на гетерозиготность изученной коротколистной материнской особи и достаточно сложный тип наследования морфологии листовых пластинок.
Полученные результаты рекомендуется использовать при описании формового разнообразия ивы трехтычинковой в природных популяциях, а также при планировании селекционного процесса и оценке полученного материала. В частности, возможность появления новых вариантов признаков в результате апомиксиса заставляет критически отнестись к получению исходного материала путем гибридизации.
| Конфликт интересов Не указан. | Conflict of Interest None declared. |
Список литературы / References
- Skvortsov A.K. Willows of Russia and Adjacent Countries. Taxonomical and Geographical Revision / A.K. Skvortsov. – Joensuu: University of Joensuu, 1999. – 307 p.
- Bűchler W. Neue Chromosomenzählungen in der Gattung Salix / W. Bűchler // Bot. Helvet. – 1985. – N – S. 165–175.
- Сукачев В.Н. Из работ по селекции ивы / В.Н. Сукачев // Селекция и интродукция быстрорастущих древесных пород. – Л.: Гослестехиздат, 1934. – С. 51–85.
- Фучило Я.Д. Селекційні основи виведення нових сортів рослин сімейства Вербові (Salicaceae Mirb.) для створення енергетичних плантацій / Я.Д. Фучило, O.O. Афонiн, М.В. Сбитна // Сортовивчення та охорона прав на сорти рослин. – 2016. – №4. – С. 18-25.
- Wichura M. Bastardbefruchtung im Pflanzenreich, erläutert an den Bastarden der Weiden / M. Wichura. – Breslau: Verlag von E. Morgenstern, 1865. – 95 p.
- Braun A. Über Parthenogenesis bei Pflanzen / A. Braun // Physikalische Abhandlungen der Konigl. Academie der Wissenschaften zu Berlin. – Berlin, 1857. – S. 311–376.
- Winkler H. Parthenogenesis und Apogamie im Pflanzenreiche / Winkler. – Jena: Verlag von G. Fischer, 1908. – 166 S.
- Хохлов С.С. Апомиксис: классификация и распространение у Покрытосеменных растений / С.С. Хохлов // Усп. соврем. ген. – М.: Наука, 1967. – Вып. 1. – С. 43–105.
- Кашин А.С. Гаметофитный апомиксис как неустойчивая система семенного размножения у цветковых / А.С. Кашин. – Саратов: Науч. книга, 2006. – 309 с.
- Соколов В.А. Существует ли гаметофитный апомиксис у диплоидных цветковых растений? / В.А. Соколов, П.А. Панихин, Т.К. Тараканова // Вавиловский журнал генетики и селекции. – 2011. – Т.15. – №1. – С. 80–101.
- Афонин А.А. Редуцированный гаметофитный апомиксис в популяциях ивы шерстистопобеговой (Salix dasyclados Wimm. = S. gmelinii Pall.) / А.А. Афонин // Бюлл. Главного ботанического сада. – М.: Научтехлитиздат, 2018. – №1 (204). – С. 43–51.
- Угольникова Е.В. Особенности репродуктивной биологии ив (Salix L.) в Саратовской области / Е.В. Угольникова, А.С. Кашин // Ботан. журн. – 2013. – Т. 98. – № – С. 723–732.
- Kuzovkina Y.A. Compilation of the Checklist for Cultivars of Salix (Willow) / Y.A. Kuzovkina // HortScience. – 2015. – N 50. – P. 1608–1609.
- Баркалов В.Ю. Филогенетические отношения видов Salix L. subg. Salix (Salicaceae) по данным секвенирования межгенных спейсеров хлоропластного генома и ITS ядерной рибосомальной ДНК / В.Ю. Баркалов, М.М. Козыренко // Генетика. – 2014. – Т. 50. – № 8. – С. 940–949.
- Blackburn K. A preliminary account of the chromosomes and chromosome behavior in the Salicaceae / K. Blackburn, J. Harrison // Ann. Bot. – 1924. – N 38. – P. 361–378.
- Håkansson A. Chromosome Numbers and Meiosis in certain Salices / A. Håkansson // Heriditas. – 1955. – B. 41. – N 3–4. – P. 454–482.
- Wilkinsson J. The cytology of Salix in relation to its taxonomy / J. Wilkinsson // Ann. Bot. NS. – 1944. – P. 269–283.
- Neumann A. Cytotaxonomischer Beitrag zur Gattung Salix / A. Neumann, A. Polatschek // Ann. Naturhistor. Mus. – 1972. – Vol. 76. – S. 619-633.
- Hörandl E. Die Gattung Salix in Österreich mit Berücksichtigung angrenzender Gebiete / E. Hörandl // Abh. der Zool.-Bot. Ges. – 1992. – N – 170 S.
- Афонин А.А. Формовое разнообразие ивы трехтычинковой (Salix triandra L.) на территории Восточной Европы / А.А. Афонин, Я.Д. Фучило // Вестник БГУ. – 2012. – № – С. 32–36.
Список литературы на английском языке / References in English
- Skvortsov A.K. Willows of Russia and Adjacent Countries. Taxonomical and Geographical Revision / A.K. Skvortsov. – Joensuu: University of Joensuu, 1999. – 307 p.
- Bűchler W. Neue Chromosomenzählungen in der Gattung Salix [New chromosome counts in the genus Salix] / W. Bűchler // Bot. Helvet. – 1985. – N 95. – P. 165–175. [in German]
- Sukachev V.N. Iz rabot po selektsii ivy [From Works on Willow Breeding] / V.N. Sukachev // Selektsiya i introduktsiya bystrorastushchikh drevesnykh porod [Selection and Introduction of Fast-growing Tree Species]. – L.: Goslestekhizdat, 1934. – P. 51-85. [in Ukrainian]
- Fuchilo Ya.D. Selektsіinі osnovi vivedennya novikh sortіv roslyn sіmeistva Verbovі (Salicaceae Mirb.) dlia stvorennya energetichnikh plantatsіi [Selection Bases for Emergence of New Varieties of Plants of Family Verbia (Salicaceae Mirb.) for Creation of Energy Plantations] / Ya.D. Fuchilo, O.O. Afonin, M.V. Sbytna // Sortovyvchennya ta okhorona prav na sorty roslyn [Sorting and Protecting Rights to Plant Varieties]. – 2016 – No.4. – P. 18-25. [in Ukrainian]
- Wichura M. Bastardbefruchtung im Pflanzenreich, erläutert an den Bastarden der Weiden [Bastardbefrauen in the plant kingdom, explained at the bastards of the pastures] / M. Wichura. – Breslau: Verlag von E. Morgenstern, 1865. – 95 p. [in German]
- Braun A. Über Parthenogenesis bei Pflanzen [About parthenogenesis in plants] / A. Braun // Physikalische Abhandlungen der Konigl. Academie der Wissenschaften zu Berlin. [Physical essays by Konigl. Academy of Sciences in Berlin] – Berlin, 1857. – P. 311–376. [in German]
- Winkler H. Parthenogenesis und Apogamie im Pflanzenreiche [Parthenogenesis and apogamy in the vegetable kingdom] / Winkler. – Jena: Verlag von G. Fischer, 1908. – 166 P. [in German]
- Khokhlov S.S. Apomiksis: klassifikaciya i rasprostranenie u Pokrytosemennykh rastenii [Apomixis: Classification and Distribution of Metasperm] / S.S. Khokhlov // sovrem. gen [Success of Modern Gen.] – M.: Nauka, 1967. – Is. 1 – P. 43-105. [in Russian]
- Kashin A.S. Gametofitnyi apomiksis kak neustoichivaya sistema semennogo razmnozheniya u tsvetkovykh [Gametophytic Apomixis as Unstable System of Seed Reproduction in Flowering Plants] / A.S. Kashin. - Saratov: Sci. Book, 2006. - 309 p. [in Russian]
- Sokolov V.A. Sushhestvuet li gametofitnyi apomiksis u diploidnykh tsvetkovykh rastenii? [Is there a Gametophytic Apomixis in Diploid Flowering Plants?] / V.A. Sokolov, P.A. Panikhin, T.K. Tarakanova // Vavilovskiy zhurnal genetiki i selektsii [Vavilov Journal of Genetics and Selection] – 2011. – V.15. - No.1. – P. 80-101. [in Russian]
- Afonin A.A. Redutsirovannyi gametofitnyi apomiksis v populyatsiyakh ivy sherstistopobegovoi (Salix dasyclados = S. gmelinii Pall.) [Reduced Gametophytic Apomixis in Populations of Willow (Salix dasyclados Wimm. = S. gmelinii Pall)] / A.A. Afonin // Byull. Glavnogo botanicheskogo sada [Bull. of Main Botanical Garden]. - M.: Nauchtekhlizdat, 2018. - No. 1 (204). - P. 43-51. [in Russian]
- Ugolnikova E.V. Osobennosti reproduktivnoi biologii iv (Salix) v Saratovskoi oblasti [Features of Reproductive Biology (Salix L.) in the Saratov Region] / E.V. Ugolnikova, A.S. Kashin // Botan. Journ [Botan. Journ] - 2013. - Vol. 98. - No.6. - P. 723-732. [in Russian]
- Kuzovkina Y.A. Compilation of the Checklist for Cultivars of Salix (Willow) / Y.A. Kuzovkina // HortScience. – 2015. – N 50. – P. 1608–1609.
- Barkalov V.Yu. Filogeneticheskie otnosheniya vidov Salix subg. Salix (Salicaceae) po dannym sekvenirovaniya mezhgennykh speiserov khloroplastnogo genoma i ITS yadernoi ribosomalnoi DNK [Phylogenetic Relations of Species Salix L. subg. Salix (Salicaceae) according to Sequencing of Intergenic Spacers of Chloroplast Genome and ITS Nuclear Ribosomal DNA] / V.Yu. Barkalov, M.M. Kozyrenko // Genetika [Genetics]. – 2014. – Vol. 50. – No.8. – P. 940-949. [in Russian]
- Blackburn K. A preliminary account of the chromosomes and chromosome behavior in the Salicaceae / K. Blackburn, J. Harrison // Ann. Bot. – 1924. – N 38. – P. 361–378.
- Hakansson A. Chromosome Numbers and Meiosis in certain Salices / A. Hakansson // Heriditas. – 1955. – B. 41. – N 3–4. – P. 454–482.
- Wilkinsson J. The cytology of Salix in relation to its taxonomy / J. Wilkinsson // Ann. Bot. NS. – 1944. – P. 269–283.
- Neumann A. Cytotaxonomischer Beitrag zur Gattung Salix [Cytotaxonomic contribution to the genus Salix] / A. Neumann, A. Polatschek // Ann. Naturhistor. Mus. – 1972. – Vol. 76. – P. 619-633. [in German]
- Hörandl E. Die Gattung Salix in Österreich mit Berücksichtigung angrenzender Gebiete [The genus Salix in Austria with consideration of adjacent areas] / E. Hörandl // Abh. der Zool.-Bot. Ges. – 1992. – N 27. – 170 p. [in German]
- Afonin A.A. Formovoe raznoobrazie ivy trekhtychinkovoi (Salix triandra) na territorii Vostochnoi Evropy [Formative Variety of Almpnd-Leaved Willow (Salix triandra L.) in Eastern Europe] / A.A. Afonin, Ya.D. Fuchilo // Vestnik BGU [Bulletin of BSU]. – 2012. – No.4. – P. 32-36. [in Russian]