Molecular Genetic Characterization of Brown Cattle Bred on the Territory of the Republic of Dagestan
Molecular Genetic Characterization of Brown Cattle Bred on the Territory of the Republic of Dagestan
Abstract
The necessity to preserve the gene pool of the mountain cattle and Caucasian brown breed lies in the specifics of the mountain zone, where intensification of cattle breeding is complicated by natural and geographical conditions. Study of allelic diversity of these breeds using different types of DNA markers will help to evaluate the degree of inbreeding and analyse the structure of populations. A total of 424 alleles were found, of which 27 are private. The study of the structure of the analysed populations showed their high level of belonging to their own cluster, as well as the separation of Highland cattle and Caucasian brown cattle into 2 subgroups, which is of interest in biodiversity conservation. Also, 86 sequences of mitochondrial DNA D-loop were obtained from the investigated cattle breeds. When studying the median network, a division into 5 clusters corresponding to haplogroups and subhaplogroups (P, Q, T1, T2, T3) was observed.
1. Введение
Дагестан – своеобразный и интересный край, расположенный в самой южной части Российской Федерации. Он расположен на северо-восточных склонах Большого Кавказа и Прикаспийской низменности. Почти половину территории республики (48%) занимает горная зона . Изучение трудовой деятельности населения Дагестана показывает, что источником существования горцев был тяжелый труд, который осложнялся неблагоприятными природными условиями: пониженное атмосферное давление, разреженный воздух, недостаток кислорода, резкие перепады температур (день-ночь) и сложный рельеф . Вместе с тем большие территории горных пастбищ, дешевый пастбищный корм и большая продолжительность пастбищного сезона благоприятствовали разведению и выращиванию крупного рогатого скота. В Дагестане испокон веков занимаются скотоводством и вплоть до тридцатых годов прошлого века разводили в горах и на равнине великокавказский и малокавказский скот. Такой скот разводят в некоторых районах и сегодня. При участии местного скота были выведены такие породы, как горский скот и кавказская бурая.
Горский скот Дагестана представляет собой весьма ценный генофонд: сравнительно небольшие размеры позволяют ему довольно свободно перемещаться в условиях высокогорья и использовать растительность в недоступных для скота других пород местах. Однако из-за низкой живой массы и молочной продуктивности его разведению в республике не уделяется должного внимания . В 2020 году было зарегистрировано всего 650 голов горского скота, находящихся в единственном генофондном хозяйстве.
Кавказская бурая порода несколько неоднородна по своему телосложению и продуктивности. Это связано с различиями между местным скотом в разных районах, степенью скрещивания со швицкой бурой, различными природно-климатическими условиями и кормлением . Для совершенствования местного скота в Дагестан завозили сперму швицких быков американской селекции, что позволило вывести новый тип бурой швицкой породы – кавказское отродье.
Одна из главных причин необходимости сохранения генофонда как горского скота, так и кавказской бурой, заключается в особенностях горной зоны, где интенсификация скотоводства затруднена природно-географическими условиями. Разведение скота узкоспециализированных заводских пород в горной зоне связано со значительной потерей их продуктивности и преждевременной выбраковкой по различным причинам . Анализ генетического разнообразия с использованием различных типов ДНК-маркеров позволит оценить аллельное разнообразие, уровень гетерозиготности и степень инбридинга, а также изучить структуру популяций этих пород скота.
Цель нашего исследования – изучение аллельного разнообразия горского скота, кавказской бурой и бурой швицкой (кавказское отродье) пород для оценки степени инбридинга и анализа структуры популяций с использованием различных типов ДНК-маркеров (STR-маркеры и мтДНК).
2. Методы и принципы исследования
Работа была выполнена в лаборатории функциональной и эволюционной геномики животных ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста с использованием оборудования Центра коллективного пользования научным оборудованием «Биоресурсы и биоинженерия сельскохозяйственных животных» ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста. В исследованиях были использованы образцы (фрагмент уха, кровь, сперма) из коллекции ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста. Выборка включала 464 головы крупного рогатого скота, отобранные с учетом фракционального членства каждой особи в кластере на основе значений Q-критерия (коэффициента подобия), в качестве порогового значения которого выбран 50% уровень исключения. В качестве группы сравнения были взяты профили голштинской породы (табл.1).
Таблица 1 - Выборка крупного рогатого скота
Обозначение групп | Кол-во голов | Расшифровка |
DM | 131 | Горский скот, Республика Дагестан |
CB_D | 105 | Кавказская бурая, Республика Дагестан |
BS_D | 50 | Бурая швицкая, кавказское отродье, Республика Дагестан |
BS | 86 | Бурая швицкая, отечественная и зарубежная селекция |
HOL | 92 | Голштинская, Holstein Association USA, Inc. |
Выделение ДНК производилось с помощью наборов ДНК-Экстран (ЗАО «Синтол», Россия) и «COrDIS ЭКСТРАКТ» (ООО «Гордиз», Россия) в соответствии с инструкцией производителей. Качество ДНК оценивали путём электрофореза в агарозном геле. Оценка соответствия полученного раствора ДНК для секвенирования мтДНК проводилась измерением концентрации на флуориметре Qubit 3.0 и установлением соотношения А260/А280 на спектрофотометре NanоDrop8000.
ПЦР-анализ выполняли согласно «Методическим рекомендациям…» . Полиморфизм 11 STR-локусов, в том числе: TGLA227, BM2113, TGLA53, ETH10, SPS115, TGLA122, INRA23, TGLA126, BM1818, ETH225 и BM1824, рекомендованных ISAG для проведения популяционно-генетических исследований крупного рогатого скота, оценивали на 16-канальном капиллярном генетическом анализаторе ABI3130×l (Applied Biosystems, США). Исходные данные о длине аллелей были получены в программном обеспечении Gene Mapper v.4 (Applied Biosystems, США).
Амплификация и секвенирование D-петли мтДНК производились по отработанной методике с использованием коммерческих наборов в соответствии с рекомендациями производителя.
Расчет показателей, характеризующих состояние аллелофонда, и параметров генетического разнообразия производился в программе GenAIEx 6.503 . Степень генетической дифференциации между породами оценивали по показателю Fst и значениям генетических дистанций по М. Нею (DN) , . Генетические связи между изучаемыми группами были визуализированы с помощью сетей Neighbour Net на основе матрицы генетических дистанций DN в программе SplitsTree 4.14.5 . Структуру изучаемых популяций оценивали с помощью кластерного анализа в Structure 2.3.4 , использовав следующие параметры: burnin период 10000, число повторений цепей Маркова методом Монте-Карло (Markov Chain Monte Carlo, MCMC) составляло 100000 для каждого запуска. Для каждого К (от 1 до 7) проводили 10 симуляций. Программа CLUMPAK , использовалась для визуализации результатов, полученных в Structure.2.3.4, и определения наиболее вероятного числа кластеров в исследованной выборке на основании значений DeltaK по методу, предложенному Evanno et al. . Кроме того, при определении структуры популяции дополнительно учитывались полученные в CLUMPAK средние оценки сходства нескольких независимых запусков при одном значении K.
Анализ последовательностей D-петли митохондриальной ДНК проводился с использованием программ Mega7, для определения гаплотипической принадлежности использовалась программа MitoTool 1.1.2, визуализация результатов в форме медианной сети выполнялась в программе popART 1.7.
3. Основные результаты
В общей сложности для 11 проанализированных локусов было обнаружено 424 аллеля, из которых 27 были приватными (табл.2). В локусе BM1818 приватные аллели не обнаружены. Количество аллелей на локус варьировало от 22 (BM1824) до 60 (TGLA53). Горский скот выделяется наибольшим генетическим разнообразием и количеством приватных аллелей.
Таблица 2 - Генетическое разнообразие в изучаемым популяциям
Локус | Породы | В общем | ||||
DM | CB_D | BS_D | BS | HOL | ||
TGLA227 | 12 / 1 | 9 | 11 | 10 | 9 / 1 | 51 / 2 |
BM2113 | 10 / 3 | 7 | 8 / 1 | 7 | 5 | 37 / 4 |
TGLA53 | 17 / 1 | 12 | 10 | 10 | 11 | 60 / 1 |
ETH10 | 8 | 7 | 6 / 1 | 4 | 6 | 31 / 1 |
SPS115 | 9 / 1 | 7 | 6 | 5 | 6 | 33 / 1 |
TGLA122 | 17 / 4 | 10 | 7 | 11 / 2 | 7 / 2 | 52 / 8 |
INRA23 | 10 | 10 / 1 | 8 | 9 / 1 | 5 | 42 / 2 |
TGLA126 | 8 / 3 | 4 | 5 | 4 | 5 | 26 / 3 |
BM1818 | 8 | 5 | 7 | 6 | 6 | 32 |
ETH225 | 11 / 2 | 7 | 8 / 1 | 6 | 6 | 38 / 3 |
BM1824 | 6 / 2 | 4 | 4 | 4 | 4 | 22 / 2 |
| 116 / 17 | 82 / 1 | 80 / 3 | 76 / 3 | 70 / 3 | 424 / 27 |
Примечание: количество аллелей / количество приватных аллелей
В таблице 3 представлены основные статистические показатели, которые позволяют оценить состояние аллелофонда и уровень генетического разнообразия изучаемых популяций крупного рогатого скота.
Таблица 3 - Характеристика аллелофонда и генетического разнообразия изучаемых популяций крупного рогатого скота
Порода | n | Na | Ne | Na >= 5% | Ho | He | Fis |
DM | 131 | 10,55±1,08 | 5,00±0,64 | 4,91±0,34 | 0,72±0,04 | 0,76±0,03 | 0,053 |
CB_D | 105 | 7,45±0,78 | 3,74±0,48 | 4,18±0,38 | 0,75±0,04 | 0,70±0,03 | -0,066 |
BS_D | 50 | 7,27±0,62 | 3,87±0,39 | 4,73±0,43 | 0,76±0,04 | 0,71±0,03 | -0,062 |
BS | 86 | 6,91±0,80 | 3,77±0,34 | 4,45±0,37 | 0,72±0,04 | 0,71±0,03 | -0,010 |
HOL | 92 | 6,36±0,61 | 3,60±0,38 | 4,00±0,45 | 0,68±0,04 | 0,69±0,03 | 0,018 |
Примечание: n – количество голов в выборке; Na – среднее число аллелей на локус; Ne – эффективное число аллелей на локус; Na >= 5% - число информативных аллелей, т.е. встречающиеся с частотой от 5% и выше; Ho – наблюдаемая гетерозиготность; He – ожидаемая гетерозиготность; Fis – индекс фиксации
Наименьшим числом аллелей в расчете на 1 локус характеризовалась группа голштинского скота, в то время как горский скот – наибольшим. Эффективное число аллелей варьировало от 3,60 в группе голштинского скота до 5,00 в группе горского скота. Для горского скота также было идентифицировано и максимальное число информативных аллелей. Анализ параметров генетического разнообразия показал, что у всех групп, за исключением группы голштинского скота, уровень наблюдаемой гетерозиготности превышал 0,70. Рассматривая значение индекса фиксации, во всех группах, кроме горского и голштинского скота, наблюдается избыток гетерозигот, при этом обнаруженный высокий дефицит гетерозигот у горского скота (0,053) может быть вызван низкой численностью популяции.
Анализ структуры генетической сети (рис.1), отображающей взаимосвязи изучаемых популяций показал, что, хотя бурые породы и сформировали единый кластер, значительно удаленный от группы сравнения, голштинской породы, каждая из исследованных популяций представляет собой достаточно обособленную группу, отличающуюся от других бурых пород.
Исследование структуры изучаемых популяций (рис.2) показало их высокий уровень принадлежности к собственному кластеру. При К=4 образцы животных кавказского отродья бурой швицкой породы образовали единый кластер с чистопородными животными бурой швицкой породы, но при дальнейшем увеличении К до 7 выделились в собственный кластер. При этом же значении К, горский скот и кавказская бурая разделяются на 2 подгруппы, что представляет интерес при сохранении биоразнообразия. Также были выявлены животные как среди горского скота, так и среди бурой швицкой, внесших свой в клад в аллельное разнообразие кавказской бурой породы и кавказского отродья бурой швицкой породы.
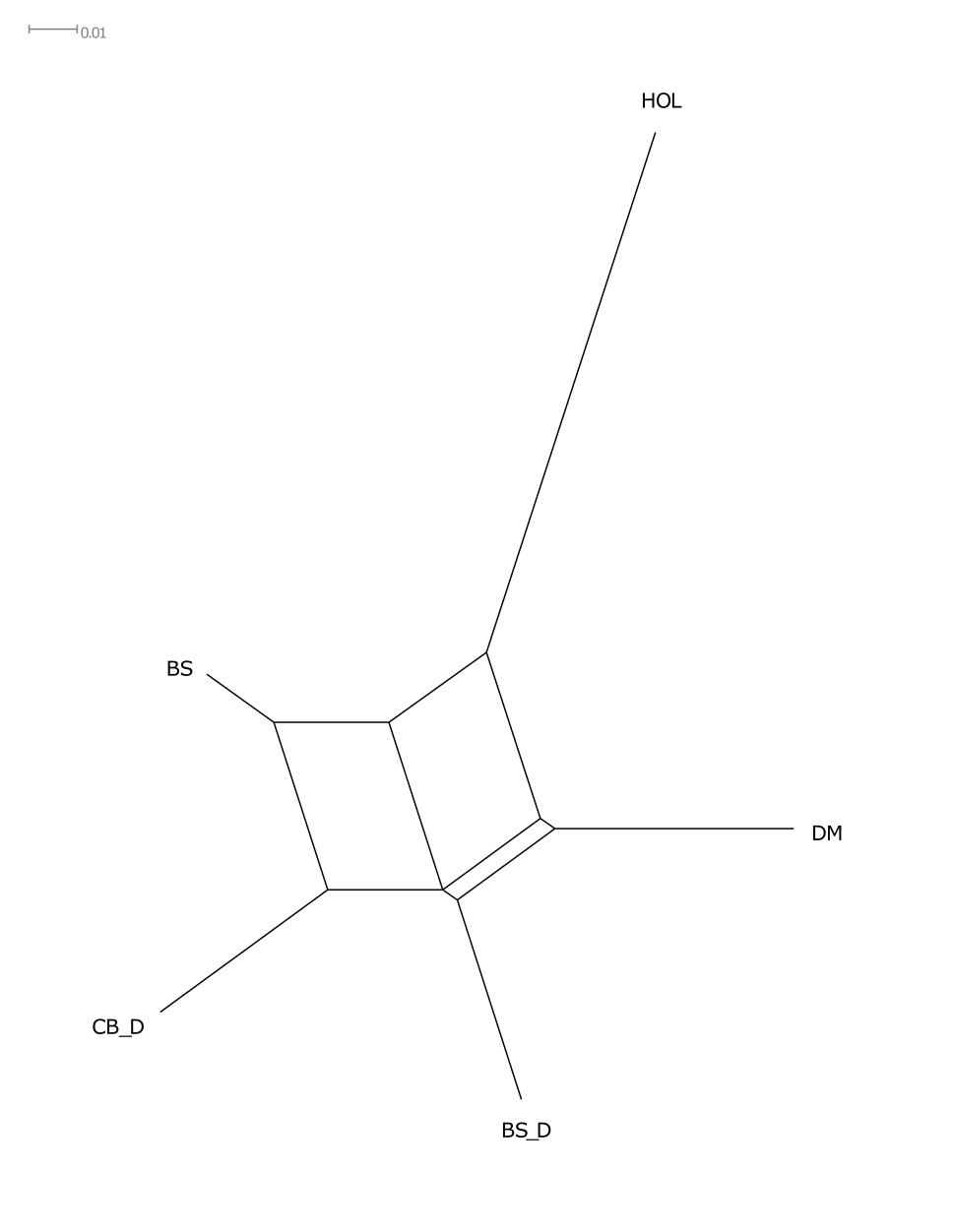
Рисунок 1 - Дендрограмма на основании попарных генетических дистанций (DJost), построенная по алгоритму NeighborNet
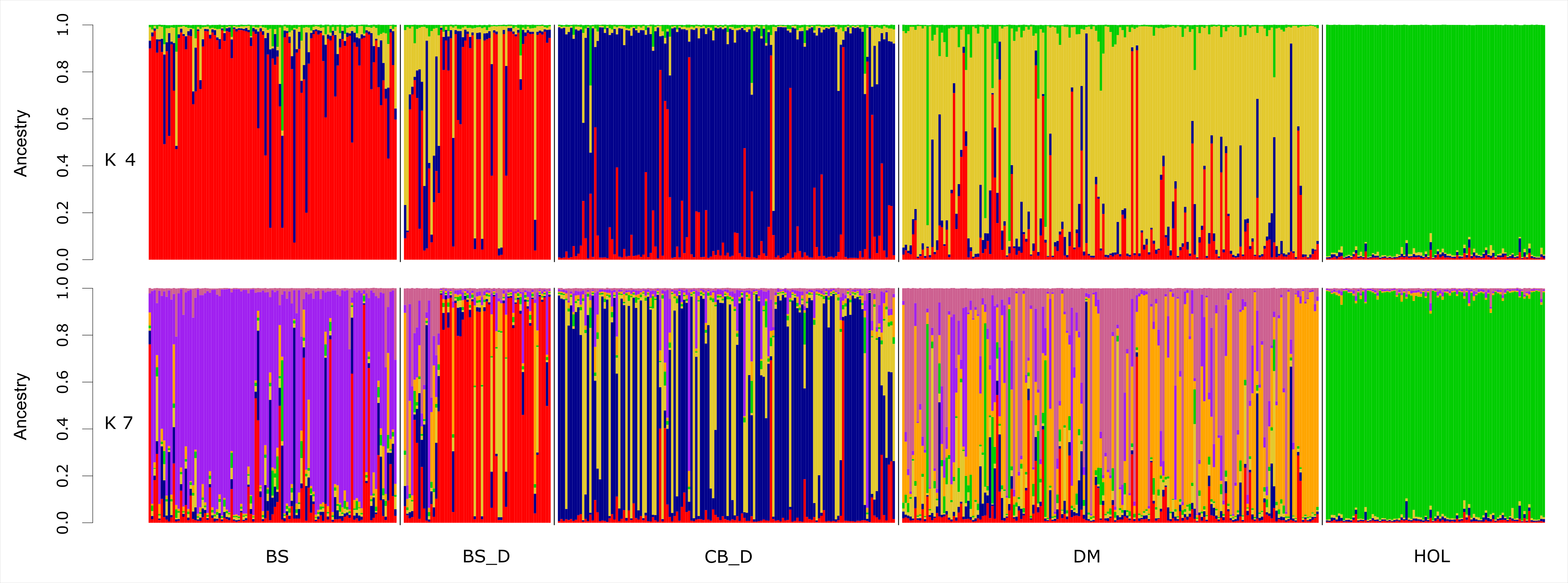
Рисунок 2 - Структура популяций исследуемых пород крупного рогатого скота
Основными параметрами, характеризующими разнообразие популяций на основании анализа полиморфизма митохондриальной ДНК, является среднее число нуклеотидных различий (К), гаплотипическое разнообразие (HD) и нуклеотидное разнообразие (π). HD – вероятность того, что два случайно выбранных гаплотипа различны , π – среднее число нуклеотидных различий на сайт между двумя случайно выбранными последовательностями . Показатели генетического разнообразия в изученных популяциях в целом обобщены в таблице 4.
Всего было получено 86 последовательностей D-петли митохондриальной ДНК от изучаемых пород крупного рогатого скота. Последовательности D-петли длиной 898 п.о. (выровненные позиции от 15791 до 16338 на референсный полный митохондриальный геном крупного рогатого скота Bos taurus mitochondrion, complete genome NC_006853.1) характеризовались наличием 50 гаплотипов, обусловленных 86 полиморфными сайтами. При этом 33 гаплотипа оказались уникальными, т.е. присутствовали только у одного животного.
Таблица 4 - Показатели генетического разнообразия у изучаемых пород на основании анализа последовательности D-петли мтДНК
Pop | n | S | K | H | Hd (±SD) | π (±SD) | Tajima’s D | Fu’s Fs |
DM | 25 | 36 | 6,770 | 17 | 0,963±0,021 | 0,00756± 0,00075 | 1,17170 ns | -5,014* |
CB_D | 22 | 29 | 5,870 | 14 | 0,961±0,022 | 0,00654±0,00057 | -1,00394 ns | -3,434 ns |
BS_D | 22 | 41 | 6,268 | 17 | 0,965±0,028 | 0,00698±0,00157 | -1,78475 ns | -7,055** |
BS | 17 | 19 | 3,221 | 12 | 0,956±0,033 | 0,00359±0,00043 | -1,68225 ns | -6,003** |
Примечание: n – количество образцов; S – число полиморфных сайтов; H – число гаплотипов; HD – гаплотипическое разнообразие; K – среднее число нуклеотидных различий; π – нуклеотидное разнообразие; SD – стандартное отклонение; Tajima's D и Fu's Fs – индексы селективной нейтральности [17,18]; ns – не достоверно; * – P < 0.05, ** – P < 0.01, *** – P < 0.001
Все исследуемые породы крупного рогатого скота характеризовались высоким уровнем гаплотипического разнообразия (Hd), которое варьировало от 0,956±0,033 в кавказском отродье бурой швицкой породы до 0,965±0,028 у чистопородной бурой швицкой. В кавказском отродье бурой швицкой породы и горском скоте было идентифицировано наибольшее количество гаплотипов (H=17), в то время как наименьшим количеством характеризовалась чистопородная бурая швицкая. К тому же у нее было обнаружено самое низкое значение нуклеотидного разнообразия (π=0,00359±0,00043). В других породах разнообразие нуклеотидов было значительно выше и варьировало от 0,00654±0,00057 в кавказской бурой породе до 0,00756±0,00075 у горского скота.
Высокий уровень гаплотипического и нуклеотидного разнообразия, обнаруженные в породах кавказская бурая, кавказском отродье бурой швицкой породы и горском скоте, свидетельствует о наличии в этих группах большого количества семейств различного происхождения. Тогда как высокие показатели гаплотипического разнообразия на фоне более низкого нуклеотидного разнообразия, обнаруженные у чистопородной бурой швицкой породы, указывают на присутствие большого числа близко родственных гаплотипов, что может быть следствием развития данной группы животных с использованием небольшого количества основателей.
Во всех исследуемых группах крупного рогатого скота были обнаружены отрицательные значения индексов нейтральности Tajima's D и Fu's Fs, кроме группы горского скота, где индекс Tajima's D имел незначимое положительное значение (Tajima's D=1,17170 ns). При этом, более чувствительный показатель Fu's Fs в этой группе имел достоверное отрицательное значение (Fu's Fs=5,014), что говорит об экспансивном росте численности данной популяции. Другие группы КРС также характеризовались достоверным отрицательным значением индекса Fu's Fs, который указывает на избыток редких мутаций в популяциях, что подтверждает экспансивный рост. Однако, кавказская бурая порода крупного рогатого скота характеризовалась незначимыми отрицательными значения обоих критериев, что может указывать лишь на тенденцию увеличения численности данной популяции.
В структуре медианной сети (рис.3) наблюдается разделение на 5 кластеров, соответствующих гаплогруппам и подгаплогруппам (P, Q, T1, T2, T3). Животные разных групп кластеризовались в различные кластеры, кроме чистопородной бурой швицкой породы. Все животные этой группы соотносились к гаплогруппе T3. Такое высокое гаплогрупповое разнообразие обусловлено большим количеством приватных гаплотипов в исследуемой выборке животных.
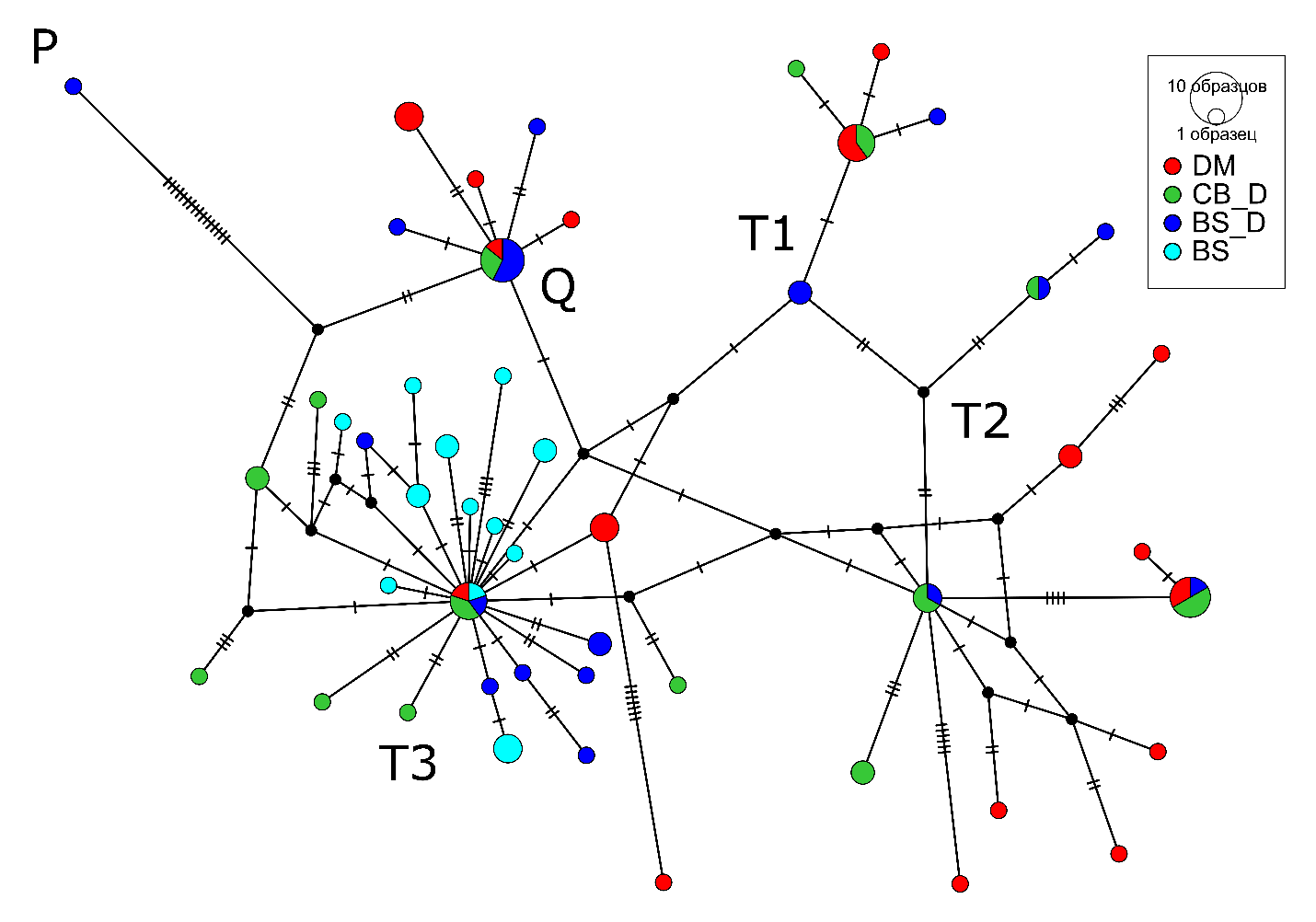
Рисунок 3 - Медианная сеть гаплотипов мтДНК исследуемых пород крупного рогатого скота
Таблица 5 - Распределение по гаплогруппам у изучаемых пород на основании анализа последовательности D-петли мтДНК
Порода | Гаплогруппа | |||||||||
P | Q | T1 | T2 | T3 | ||||||
кол-во | % | кол-во | % | кол-во | % | кол-во | % | кол-во | % | |
DM | - | - | 6 | 24 | 4 | 16 | 10 | 40 | 5 | 20 |
CB_D | - | - | 2 | 9,09 | 3 | 13,64 | 8 | 36,36 | 9 | 40,91 |
BS_D | 1 | 4,55 | 6 | 27,27 | 3 | 13,64 | 4 | 18,18 | 8 | 36,36 |
BS | - | - | - | - | - | - | - | - | 17 | 100 |
4. Обсуждение
В Дагестане испокон веков занимаются скотоводством, но низкая молочная и мясная продуктивность местного скота не могли обеспечить растущие потребности населения в продукции животноводства. Известные исследователи крупного рогатого скота Кавказа А.А. Калантар (1890, 1901), А. Браунер (1933), А.З. Тамамшев (1923, 1947) на основании краниологических и других данных пришли к заключению, что между кавказским скотом и древним торфянниковым, который является родоначальником швицкого скота, есть определенное сходство, что в какой-то степени, явилось предпосылкой для использования швицких быков с целью улучшения продуктивных качеств аборигенного скота на Кавказе .
Полученные нами данные по STR-маркерам схожи с данными, полученными в более ранних исследованиях , однако, некоторое снижение аллельного разнообразия и уменьшение приватных аллелей у горского скота связано с уменьшением числа животных в выборке на основании выбраковки помесных животных.
Анализ по мтДНК показал, что бурая швицкая порода, а также наибольший процент кавказской бурой и кавказского отродья бурой швицкой, относятся к широко распространенной европейской гаплогруппе Т3. Тогда как другие выявленные гаплогруппы сосредоточены: T1 – в Африке, а T2 – на Ближнем Востоке. Редкая гаплогруппа Q имеет ближневосточное происхождение, а P – была характерна для тура (Bos primigenius), прародителя современного крупного рогатого скота , , , . Все животные горского скота с гаплогруппой Q происходили из отдалённого горного района . Можно предположить, что предки животных группы CB_D и BS_D, с той же гаплогруппой, происходят из этого же отдаленного региона.
5. Заключение
Таким образом, проведенные нами исследования представляют собой наиболее полные сведения о состоянии аллелофонда, генетическом разнообразии и дифференциации локальных пород Дагестана. Сохранение ценного генофонда этих пород является необходимым для сельского хозяйства горной зоны Дагестана, а также создания высокопродуктивных животных, хорошо приспособленных к суровым условиям горной зоны.